Часть I Общая физиология
Любому тренеру и педагогу для успешной профессиональной деятельности необходимо знание функций организма человека. Лишь учет особенностей его жизнедеятельности может помочь правильно управлять ростом и развитием организма человека, сохранением здоровья детей и взрослых, поддержанием работоспособности даже в пожилом возрасте, рациональному использованию мышечных нагрузок в процессе физического воспитания и спортивной тренировки.
1. Введение. История физиологии
Датой становления современной физиологии является 1628 г., когда английский врач и физиолог Вильям Гарвей опубликовал результаты своего исследования по кровообращению у животных.
Физиология – наука о функциях и механизмах деятельности клеток, тканей, органов, систем и всего организма в целом. Физиологической функцией является проявление жизнедеятельности организма, имеющее приспособительное значение.
1.1. Предмет физиологии, ее связь с другими науками и значение для физической культуры и спорта
Физиология как наука неразрывно связана с другими дисциплинами. Она базируется на знаниях физики, биофизики и биомеханики, химии и биохимии, общей биологии, генетики, гистологии, кибернетики, анатомии. В свою очередь, физиология является основой медицины, психологии, педагогики, социологии, теории и методики физического воспитания. В процессе развития физиологической науки из общей физиологии выделились различные частные разделы: физиология труда, физиология спорта, авиакосмическая физиология, физиология подводного труда, возрастная физиология, психофизиология и др.
Общая физиология представляет собой теоретическую основу физиологии спорта. Она описывает основные закономерности деятельности организма людей разного возраста и пола, различные функциональные состояния, механизмы работы отдельных органов и систем организма и их взаимодействия. Ее практическое значение состоит в научном обосновании возрастных этапов развития организма человека, индивидуальных особенностях отдельных людей, механизмов проявления их физических и умственных способностей, особенностей контроля и возможностей управления функциональным состоянием организма. Физиология вскрывает последствия вредных привычек у человека, обосновывает пути профилактики функциональных нарушений и сохранение здоровья. Знания физиологии помогают педагогу и тренеру в процессах спортивного отбора и спортивной ориентации, в прогнозировании успешности соревновательной деятельности спортсмена, в рациональном построении тренировочного процесса, в обеспечении индивидуализации физических нагрузок и открывают возможности использования функциональных резервов организма.
1.2. Методы физиологических исследований
Физиология – наука экспериментальная. Знания о функциях и механизмах деятельности организма построены на опытах, проводимых на животных, наблюдениях в клинике, обследованиях здоровых людей в различных экспериментальных условиях. При этом в отношении здорового человека требуются методы, не связанные с повреждениями его тканей и проникновением во внутрь организма – так называемые неинвазивные методы.
В общей форме физиология использует три методических приема исследований: наблюдение, или метод «черного ящика», острый опыт и хронический эксперимент.
Классическими методами исследований являлись методы удаления и методы раздражения отдельных частей или целых органов, в основном применявшиеся в опытах на животных или во время операций в клинике. Они давали приблизительное представление о функциях удаленных или раздражаемых органов и тканей организма. В этом отношении прогрессивным методом исследования целостного организма стал метод условных рефлексов, разработанный И. П. Павловым.
В современных условиях наиболее распространены электрофизиологические методы, позволяющие регистрировать электрические процессы, не изменяя текущей деятельности изучаемых органов и без повреждения покровных тканей, – например, электрокардиография, электромиография, электроэнцефалография (регистрация электрической активности сердца, мышц и мозга). Развитие радиотелеметрии позволяет передавать эти получаемые записи на значительные расстояния, а компьютерные технологии и специальные программы обеспечивают тонкий анализ физиологических данных. Использование фотосъемки в инфракрасных лучах (тепловидения) позволяет выявить наиболее горячие или холодные участки тела, наблюдаемые в состоянии покоя или в результате деятельности. С помощью так называемой компьютерной томографии, не вскрывая мозга, можно увидеть морфофункциональные его изменения на различной глубине. Новые данные о работе мозга и отдельных частей тела дает изучение магнитных колебаний.
1.3. Краткая история физиологии
Наблюдения за жизнедеятельностью организма производились с незапамятных времен. В XIV–XV веках до н. э. в Древнем Египте при изготовлении мумий люди хорошо знакомились с внутренними органами человека. В гробнице врача фараона Унаса изображены древние медицинские инструменты. В Древнем Китае только по пульсу удивительно тонко различали до 400 болезней. В IV–V веке до н. э. там было развито учение о функционально важных точках тела, которое в настоящее время явилось основой для современных разработок рефлексотерапии и иглоукалывания, Су-Джок терапии, тестирования функционального состояния скелетных мышц спортсмена по величине напряженности электрического поля кожи в биоэлектрически активных точках над ними. Древняя Индия прославилась своими особыми растительными рецептами, воздействием на организм упражнениями йоги и дыхательной гимнастики. В Древней Греции первые представления о функциях мозга и сердца высказывали в IV–V веке до н. э. Гиппократ (460–377 до н. э.) и Аристотель (384–322 до н. э.), а в Древнем Риме во II веке до н. э. – врач Гален (201–131 до н. э.).
Как экспериментальная наука физиология возникла в XVII веке, когда английский врач В. Гарвей открыл круги кровообращения. В этот же период французский ученый Р. Декарт ввел понятие рефлекс (отражение), описав путь внешней информации в мозг и обратный путь двигательного ответа. Работами гениального русского ученого М. В. Ломоносова и немецкого физика Г. Гельмгольца о трехкомпонентной природе цветного зрения, трактатом чеха Г. Прохазки о функциях нервной системы и наблюдениями итальянца Л. Гальвани о животном электричестве в нервах и мышцах отмечен XVIII век. В XIX веке разработаны представления английского физиолога Ч. Шеррингтона об интегративных процессах в нервной системе, изложенные в его известной монографии в 1906 г. Проведены первые исследования утомления итальянцем А. Моссо. Обнаружил изменения постоянных потенциалов кожи при раздражениях у человека И. Р. Тарханов (феномен Тарханова).
В XIX в. работами «отца русской физиологии» И. М. Сеченова (1829–1905) заложены основы развития многих областей физиологии – изучение газов крови, процессов утомления и «активного отдыха», а главное – открытие в 1862 г. торможения в центральной нервной системе («Сеченовского торможения») и разработка физиологических основ психических процессов человека, показавших рефлекторную природу поведенческих реакций человека («Рефлексы головного мозга», 1863). Дальнейшая разработка идей И. М. Сеченова шла двумя путями. С одной стороны, изучение тонких механизмов возбуждения и торможения проводилось в Санкт-Петербургском университете И. Е. Введенским (1852–1922). Им создано представление о физиологической лабильности как скоростной характеристике возбуждения и учение о парабиозе как общей реакции нервно-мышечной ткани на раздражение. В дальнейшем это направление было продолжено его учеником А. А. Ухтомским (1875–1942), который, изучая процессы координации в нервной системе, открыл явление доминанты (господствующего очага возбуждения) и роль в этих процессах усвоения ритма раздражений. С другой стороны, в условиях хронического эксперимента на целостном организме И. П. Павлов (1849–1936) впервые создал учение об условных рефлексах и разработал новую главу физиологии – физиологию высшей нервной деятельности. Кроме того, в 1904 г. за свои работы в области пищеварения И. П. Павлов, одним из первых русских ученых, был отмечен Нобелевской премией. Физиологические основы поведения человека, роль сочетанных рефлексов были разработаны В. М. Бехтеревым.
Крупный вклад в развитие физиологии внесли и другие выдающиеся отечественные физиологи: основатель эволюционной физиологии и адаптологии академик Л. А. Орбели; изучавший условно-рефлекторные влияния коры на внутренние органы акад. К. М. Быков; создатель учения о функциональной системе акад. П. К. Анохин; основатель отечественной электроэнцефалографии акад. М. Н. Ливанов; разработчик космической физиологии – акад. В. В. Парин; основатель физиологии активности Н. А. Бернштейн и многие др.
В области физиологии мышечной деятельности следует отметить основателя отечественной физиологии спорта – проф. А. Н. Крестовникова (1885–1955), написавшего первый учебник по физиологии человека для физкультурных вузов страны (1938) и первую монографию по физиологии спорта (1939), а также широко известных ученых – проф. Е. К. Жукова, В. С. Фарфеля, Н. В. Зимкина, А. С. Мозжухина и многих др., а среди зарубежных ученых – П. О. Астранда, А. Хилла, Р. Гранита, Р. Маргария и др.
2. Общие закономерности физиологии и ее основные понятия
Живые организмы представляют собой так называемые открытые системы (т. е. не замкнутые в себе, а неразрывно связанные с внешней средой). Они состоят из белков и нуклеиновых кислот и характеризуются способностью к авторегуляции и самовоспроизведению. Основные свойства живого организма – обмен веществ, раздражимость (возбудимость), подвижность, самовоспроизведение (размножение, наследственность) и саморегуляция (поддержание гомеостаза, приспособляемость-адаптивность).
2.1. Основные функциональные характеристики возбудимых тканей
Общим свойством всех живых тканей является раздражимость, т. е. способность под влиянием внешних воздействий изменять обмен веществ и энергии. Среди всех живых тканей организма особо выделяют возбудимые ткани (нервную, мышечную и железистую), реакция которых на раздражение связана с возникновением специальных форм активности – электрических потенциалов и других явлений.
Основными функциональными характеристиками возбудимых тканей являются возбудимость и лабильность.
Возбудимость – свойство возбудимых тканей отвечать на раздражение специфическим процессом возбуждения. Этот процесс включает электрические, ионные, химические и тепловые изменения, а также специфические проявления: в нервных клетках – импульсы возбуждения, в мышечных – сокращение или напряжение, в железистых – выделение определенных веществ. Он представляет собой переход из состояния физиологического покоя в деятельное состояние. Для нервной и мышечной ткани характерна также способность передавать это активное состояние соседним участкам, т. е. проводимость.
Возбудимые ткани характеризуются двумя основными нервными процессами – возбуждением и торможением. Торможение – это активная задержка процесса возбуждения. Взаимодействие этих двух процессов обеспечивает координацию нервной деятельности в целостном организме.
Различают местное (или локальное) возбуждение и распространяющееся. Местное возбуждение представляет незначительные изменения в поверхностной мембране клеток, а распространяющееся возбуждение связано с передачей всего комплекса физиологических изменений (импульса возбуждения) вдоль нервной или мышечной ткани. Для измерения возбудимости пользуются определением порога, т. е. минимальной величины раздражения, при которой возникает распространяющееся возбуждение. Величина порога зависит от функционального состояния ткани и от особенностей раздражителя, которым может быть любое изменение внешней среды (электрическое, тепловое, механическое и пр.). Чем выше порог, тем ниже возбудимость, и наоборот. Возбудимость может повышаться в процессе выполнения физических упражнений оптимальной длительности и интенсивности (например, под влиянием разминки, в ходе врабатывания) и снижаться при утомлении, развитии перетренированности.
Лабильность – скорость протекания процесса возбуждения в нервной и мышечной ткани (лат. лабилис – «подвижный»). Понятие лабильности, или функциональной подвижности, было выдвинуто Н. Е. Введенским в 1892 г. В качестве одной из мер лабильности Н. Е. Введенский предложил максимальное количество волн возбуждения (электрических потенциалов действия), которое может воспроизводиться тканью в 1 секунду в соответствии с ритмом раздражения. Лабильность характеризует скоростные свойства ткани. Она может повышаться под влиянием раздражений, тренировки, особенно у спортсменов при развитии качества быстроты.
2.2. Нервная и гуморальная регуляция функций
У простейших одноклеточных животных одна единственная клетка осуществляет разнообразные функции. Усложнение же деятельности организма в процессе эволюции привело к разделению функций различных клеток – их специализации. Для управления такими сложными многоклеточными системами уже было недостаточно древнего способа – переноса регулирующих жизнедеятельность веществ жидкими средами организма.
Регуляция различных функций у высокоорганизованных животных и человека осуществляется двумя путями: гуморальным (лат. гумор – «жидкость») – через кровь, лимфу и тканевую жидкость и нервным.
Возможности гуморальной регуляции функций ограничены тем, что она действует сравнительно медленно и не может обеспечить срочных ответов организма (быстрых движений, мгновенной реакции на экстренные раздражители). Кроме того, гуморальным путем происходит широкое вовлечение различных органов и тканей в реакцию (по принципу «Всем, всем, всем!»). В отличие от этого, с помощью нервной системы возможно быстрое и точное управление различными отделами целостного организма, доставка сообщений точному адресату. Оба эти механизма тесно связаны, однако ведущую роль в регуляции функций играет нервная система.
В регуляции функционального состояния органов и тканей принимают участие особые вещества – нейропептиды, выделяемые железой внутренней секреции гипофизом и нервными клетками спинного и головного мозга. В настоящее время известно около сотни подобных веществ, которые являются осколками белков и, не вызывая сами возбуждения клеток, могут заметно изменять их функциональное состояние. Они влияют на сон, процессы обучения и памяти, на мышечный тонус (в частности, на позную асимметрию), вызывают обездвижение или обширные судороги мышц, обладают обезболивающим и наркотическим эффектом. Оказалось, что концентрация нейропептидов в плазме крови у спортсменов может превышать средний уровень у нетренированных лиц в 6–8 раз, повышая эффективность соревновательной деятельности. В условиях чрезмерных тренировочных занятий происходит истощение нейропептидов и срыв адаптации спортсмена к физическим нагрузкам.
2.3. Рефлекторный механизм деятельности нервной системы
В деятельности нервной системы основным является рефлекторный механизм. Рефлекс – это ответная реакция организма на внешнее раздражение, осуществляемая с участием нервной системы.
Нервный путь рефлекса называется рефлекторной дугой. В состав рефлекторной дуги входят: 1) воспринимающее образование – рецептор; 2) чувствительный или афферентный нейрон, связывающий рецептор с нервными центрами; 3) промежуточные (или вставочные) нейроны нервных центров; 4) эфферентный нейрон, связывающий нервные центры с периферией; 5) рабочий орган, отвечающий на раздражение – мышца или железа.
Наиболее простые рефлекторные дуги включают всего две нервные клетки, однако множество рефлекторных дуг в организме состоят из значительного количества разнообразных нейронов, расположенных в различных отделах центральной нервной системы. Выполняя ответные реакции, нервные центры посылают команды к рабочему органу (например, скелетной мышце) через эфферентные пути, которые выполняют роль так называемых каналов прямой связи. В свою очередь, в ходе осуществления рефлекторного ответа или после него рецепторы, находящиеся в рабочем органе, и другие рецепторы тела посылают в центральную нервную систему информацию о результате действия. Афферентные пути этих сообщений – каналы обратной связи. Полученная информация используется нервными центрами для управления дальнейшими действиями, т. е. прекращением рефлекторной реакции, ее продолжением или изменением. Следовательно, основу целостной рефлекторной деятельности составляет не отдельная рефлекторная дуга, а замкнутое рефлекторное кольцо, образованное прямыми и обратными связями нервных центров с периферией.
2.4. Гомеостаз
Внутренняя среда организма, в которой живут все его клетки, – это кровь, лимфа, межтканевая жидкость. Ее характеризует относительное постоянство – гомеостаз различных показателей, так как любые ее изменения приводят к нарушению функций клеток и тканей организма, особенно высокоспециализированных клеток центральной нервной системы. К таким постоянным показателям гомеостаза относятся температура внутренних отделов тела, сохраняемая в пределах 36–37 °C, кислотно-основное равновесие крови, характеризуемое величиной рН = 7,4–7,35, осмотическое давление крови (7,6–7,8 атм), концентрация гемоглобина в крови – 130–160 г/л и др.
Гомеостаз представляет собой не статическое явление, а динамическое равновесие. Способность сохранять гомеостаз в условиях постоянного обмена веществ и значительных колебаний факторов внешней среды обеспечивается комплексом регуляторных функций организма. Эти регуляторные процессы поддержания динамического равновесия получили название гомеокинеза.
Степень сдвига показателей гомеостаза при существенных колебаниях условий внешней среды или при тяжелой работе у большинства людей очень невелика. Например, длительное изменение рН крови всего на 0,1–0,2 может привести к смертельному исходу. Однако в общей популяции имеются отдельные индивиды, обладающие способностью переносить гораздо большие сдвиги показателей внутренней среды. У высококвалифицированных спортсменов-бегунов в результате большого поступления молочной кислоты из скелетных мышц в кровь во время бега на средние и длинные дистанции рН крови может снижаться до величин 7,0 и даже 6,9. Лишь несколько человек в мире оказались способными подняться на высоту порядка 8800 м над уровнем моря (на вершину Эвереста) без кислородного прибора, т. е. существовать и двигаться в условиях крайнего недостатка кислорода в воздухе и, соответственно, в тканях организма. Эта способность определяется врожденными особенностями человека – так называемой генетической нормой реакции, которая даже для достаточно постоянных функциональных показателей организма имеет широкие индивидуальные различия.
2.5. Возникновение возбуждения и его проведение
2.5.1. Мембранные потенциалы
Мембрана клетки состоит из двойного слоя молекул липидов, повернутых «головками» наружу, а «хвостами» друг к другу. Между ними свободно плавают глыбы белковых молекул. Некоторые из них пронизывают мембрану насквозь. В части таких белков имеются особые поры или ионные каналы, через которые могут проходить ионы, участвующие в образовании мембранных потенциалов (рис. 1, А).
В возникновении и поддержании мембранного потенциала покоя основную роль играют два специальных белка. Один из них выполняет роль особого натрий-калиевого насоса, который за счет энергии АТФ активно перекачивает натрий из клетки наружу, а калий внутрь клетки. В результате концентрация ионов калия становится внутри клетки выше, чем в омывающей клетку жидкости, а ионов натрия – выше снаружи.
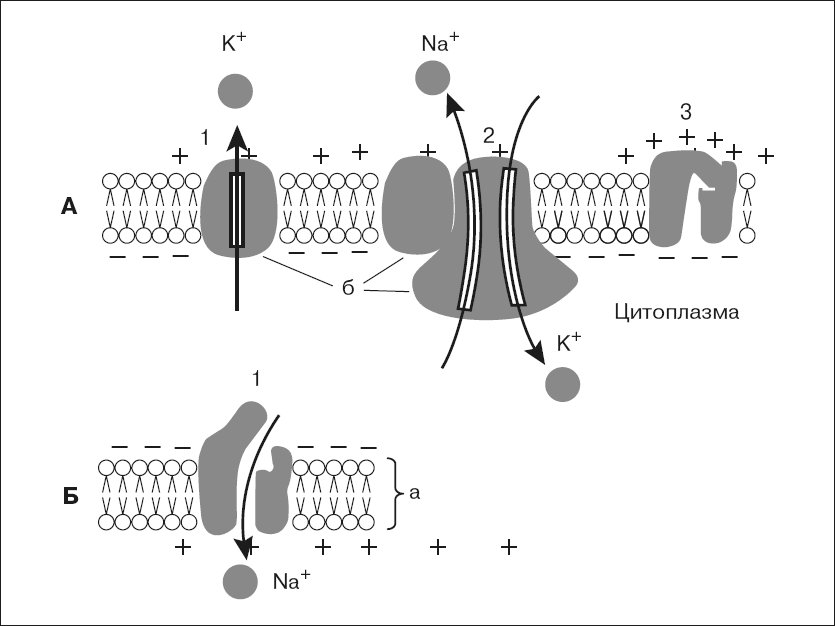
Рис. 1. Мембрана возбудимых клеток в покое (А) и при возбуждении (Б): а – двойной слой липидов, б – белки мембраны; А – каналы «утечки калия» (1), «натрий-калиевый насос» (2) и закрытый в покое натриевый канал (3); Б – открытый при возбуждении натриевый канал (1), вхождение ионов натрия в клетку и смена зарядов на наружной и внутренней стороне мембраны
Второй белок служит каналом утечки калия, через который ионы калия в силу диффузии стремятся выйти из клетки, где они содержатся в избытке. Ионы калия, выходя из клетки, создают положительный заряд на наружной поверхности мембраны. В результате внутренняя поверхность мембраны оказывается заряженной отрицательно по отношению к наружной. Таким образом, мембрана в состоянии покоя поляризована, т. е. имеется определенная разность потенциалов по обе стороны мембраны, называемая потенциалом покоя. Она равна для нейрона примерно минус 70 мВ, для мышечного волокна – минус 90 мВ. Измеряют мембранный потенциал покоя, вводя тонкий кончик микроэлектрода внутрь клетки, а второй электрод помещая в окружающую жидкость. В момент прокола мембраны и вхождения микроэлектрода внутрь клетки на экране осциллографа наблюдают смещение луча, пропорциональное величине потенциала покоя.
В основе возбуждения нервных и мышечных клеток лежит повышение проницаемости мембраны для ионов натрия – открывание натриевых каналов. Внешнее раздражение вызывает перемещение заряженных частиц внутри мембраны и уменьшение исходной разности потенциалов по обе стороны, или деполяризацию мембраны. Небольшие величины деполяризации приводят к открыванию части натриевых каналов и незначительному проникновению натрия внутрь клетки. Эти реакции являются подпороговыми и вызывают лишь местные (локальные) изменения.
При увеличении раздражения изменения мембранного потенциала достигают порога возбудимости, или критического уровня деполяризации, – около 20 мВ, при этом величина потенциала покоя снижается примерно до минус 50 мВ. В результате открывается значительная часть натриевых каналов. Происходит лавинообразное вхождение ионов натрия внутрь клетки, вызывающее резкое изменение мембранного потенциала, которое регистрируется в виде потенциала действия. Внутренняя сторона мембраны в месте возбуждения оказывается заряженной положительно, а внешняя – отрицательно (рис. 1, Б).
Весь этот процесс чрезвычайно кратковременный. Он занимает всего около 1–2 мс, после чего ворота натриевых каналов закрываются. К этому моменту достигает большой величины медленно нараставшая при возбуждении проницаемость для ионов калия. Выходящие из клетки ионы калия вызывают быстрое снижение потенциала действия. Однако окончательное восстановление исходного заряда продолжается еще некоторое время. В связи с этим в потенциале действия различают кратковременную высоковольтную часть – пик (или с пайк) и длительные малые колебания – следовые потенциалы. Потенциалы действия мотонейронов имеют амплитуду пика около 100 мВ и длительность около 1,5 мс, в скелетных мышцах – амплитуда потенциала действия 120–130 мВ, а длительность 2–3 мс.
В процессе восстановления после потенциала действия работа натрий-калиевого насоса обеспечивает «откачку» излишних ионов натрия наружу и «накачивание» потерянных ионов калия внутрь, т. е. возвращение к исходной асимметрии их концентрации по обе стороны мембраны. На работу этого механизма тратится около 70 % всей необходимой клетке энергии.
Возникновение возбуждения (потенциала действия) возможно лишь при сохранении достаточного количества ионов натрия в окружающей клетку среде. Большие потери натрия организмом (например, с потом при длительной мышечной работе в условиях высокой температуры воздуха) могут нарушить нормальную деятельность нервных и мышечных клеток, снизив работоспособность человека. В условиях кислородного голодания тканей (например, при наличии большого кислородного долга во время мышечной работы) процесс возбуждения также нарушается из-за поражения (инактивации) механизма вхождения в клетку ионов натрия, и клетка становится невозбудимой. На процесс инактивации натриевого механизма влияет концентрация ионов Са2+ в крови. При повышении содержания Са2+ снижается клеточная возбудимость, а при дефиците Са2+ возбудимость повышается и появляются непроизвольные мышечные судороги.
2.5.2. Проведение возбуждения
Потенциалы действия (импульсы возбуждения) обладают способностью распространяться вдоль по нервным и мышечным волокнам.
В нервном волокне потенциал действия является очень сильным раздражителем для соседних участков волокна. Амплитуда потенциала действия обычно в 5–6 раз превышает пороговую величину деполяризации. Это обеспечивает высокую скорость и надежность проведения.
Между зоной возбуждения (имеющей на поверхности волокна отрицательный заряд и на внутренней стороне мембраны – положительный) и соседним невозбужденным участком мембраны нервного волокна (с обратным соотношением зарядов) возникают электрические токи – так называемые местные токи. В результате развивается деполяризация соседнего участка, увеличение его ионной проницаемости и появление потенциала действия. В исходной же зоне возбуждения восстанавливается потенциал покоя. Затем возбуждением охватывается следующий участок мембраны и т. д. Таким образом с помощью местных токов происходит распространение возбуждения на соседние участки нервного волокна, т. е. проведение нервного импульса. По мере проведения амплитуда потенциала действия не уменьшается – возбуждение не затухает даже при большой длине нерва.
В процессе эволюции с переходом от безмякотных нервных волокон к мякотным произошло существенное повышение скорости проведения нервного импульса. Для безмякотных волокон характерно непрерывное проведение возбуждения, которое охватывает последовательно каждый соседний участок нерва. Мякотные же нервы почти полностью покрыты изолирующей миелиновой оболочкой. Ионные токи в них могут проходить только в оголенных участках мембраны – перехватах Ранвье, лишенных этой оболочки. При проведении нервного импульса возбуждение перескакивает от одного перехвата к другому и может включать даже несколько перехватов. Такое проведение получило название сальтаторного (лат. saltus – «прыжок»). При этом повышается не только скорость, но и экономичность проведения. Возбуждение захватывает не всю поверхность мембраны волокна, а лишь небольшую ее часть. Следовательно, меньше энергии тратится на активный транспорт ионов через мембрану при возбуждении и в процессе восстановления.
Скорость проведения в разных волокнах различна. Более толстые нервные волокна проводят возбуждение с большей скоростью: у них расстояния между перехватами Ранвье больше и длиннее скачки. Наибольшую скорость проведения имеют двигательные и проприоцептивные афферентные нервные волокна – до 100 м/с. В тонких симпатических нервных волокнах (особенно в немиелинизированных волокнах) скорость проведения мала – порядка 0,5–15 м/с.
Во время развития потенциала действия мембрана полностью теряет возбудимость. Это состояние называют полной невозбудимостью, или абсолютной рефрактерностью. За ним следует относительная рефрактерность, когда потенциал действия может возникать лишь при очень сильном раздражении. Постепенно возбудимость восстанавливается до исходного уровня.
3. Нервная система
Нервную систему подразделяют на периферическую (нервные волокна и узлы) и центральную. К центральной нервной системе (ЦНС) относят спинной и головной мозг.
3.1. Основные функции ЦНС
Все важнейшие поведенческие реакции человека осуществляются с помощью ЦНС.
Основные функции ЦНС:
• объединение всех частей организма в единое целое и их регуляция;
• управление состоянием и поведением организма в соответствии с условиями внешней среды и его потребностями.
У высших животных и человека ведущим отделом ЦНС является кора больших полушарий. Она управляет наиболее сложными функциями в жизнедеятельности человека – психическими процессами (сознание, мышление, речь, память и др.).
Основные методы изучения функций ЦНС – методы удаления и раздражения (в клинике и на животных), регистрации электрических явлений, метод условных рефлексов.
Продолжают разрабатываться новые методы изучения ЦНС: с помощью так называемой компьютерной томографии можно увидеть морфофункциональные изменения мозга на различной его глубине; фотосъемки в инфракрасных лучах (тепловидение) позволяют обнаружить наиболее «горячие» точки мозга; новые данные о работе мозга дает изучение его магнитных колебаний.
3.2. Основные функции и взаимодействия нейронов
Основными структурными элементами нервной системы являются нервные клетки или нейроны.
3.2.1. Основные функции нейронов
Через нейроны осуществляется передача информации от одного участка нервной системы к другому, обмен информацией между нервной системой и различными участками тела. В нейронах происходят сложнейшие процессы обработки информации. С их помощью формируются ответные реакции организма (рефлексы) на внешние и внутренние раздражения.
Таким образом, основными функциями нейронов являются: восприятие внешних раздражений – рецепторная функция, их переработка – интегративная функция и передача нервных влияний на другие нейроны или различные рабочие органы – эффекторная функция. В теле нервной клетки, или соме, происходят основные процессы переработки информации. Многочисленные древовидно разветвленные отростки – дендриты (греч. дендрон – «дерево») служат входами нейрона, через которые сигналы поступают в нервную клетку. Выходом нейрона является отходящий от тела клетки отросток – аксон (греч. аксис – «ось»), который передает нервные импульсы дальше – другой нервной клетке или рабочему органу (мышце, железе). Особенно высокой возбудимостью обладает начальная часть аксона и расширение в месте его выхода из тела клетки – аксонный холмик нейрона. Именно в этом сегменте клетки возникает нервный импульс.
3.2.2. Типы нейронов
Нейроны подразделяются на три основных типа: афферентные, эфферентные и промежуточные. Афферентные нейроны (чувствительные, или центростремительные) передают информацию от рецепторов в ЦНС. Тела этих нейронов расположены вне ЦНС – в спинномозговых узлах и в узлах черепных нервов. Афферентные нейроны имеют длинный отросток – дендрит, который контактирует на периферии с воспринимающим образованием – рецептором или сам образует рецептор, а также второй отросток – аксон, входящий через задние рога в спинной мозг.
Эфферентные нейроны (центробежные) связаны с передачей нисходящих влияний от вышележащих этажей нервной системы к нижележащим или из ЦНС к рабочим органам. Для эфферентных нейронов характерны разветвленная сеть коротких отростков – дендритов и один длинный отросток – аксон.
Промежуточные нейроны (интернейроны, или вставочные) – это, как правило, более мелкие клетки, осуществляющие связь между различными (в частности, афферентными и эфферентными) нейронами. Они передают нервные влияния в горизонтальном направлении (например, в пределах одного сегмента спинного мозга) и в вертикальном (например, из одного сегмента спинного мозга в другие – выше- или нижележащие сегменты). Благодаря многочисленным разветвлениям аксона промежуточные нейроны могут одновременно возбуждать большое число других нейронов.
3.2.3. Возбуждающие и тормозящие синапсы
Взаимодействие нейронов между собой (и с эффекторными органами) происходит через специальные образования – синапсы (греч. – «контакт»). Они образуются концевыми разветвлениями нейрона на теле или отростках другого нейрона. Чем больше синапсов на нервной клетке, тем больше она воспринимает различных раздражений и, следовательно, шире сфера влияний на ее деятельность и возможность участия в разнообразных реакциях организма. Особенно много синапсов в высших отделах нервной системы и именно у нейронов с наиболее сложными функциями.
В структуре синапса различают три элемента (рис. 2):
1) пресинаптическую мембрану, образованную утолщением мембраны конечной веточки аксона;
2) синаптическую щель между нейронами;
3) постсинаптическую мембрану – утолщение прилегающей поверхности следующего нейрона.
В большинстве случаев передача влияния одного нейрона на другой осуществляется химическим путем. В пресинаптической части контакта имеются синаптические пузырьки, которые содержат специальные вещества – медиаторы, или посредники. Ими могут быть ацетилхолин (в некоторых клетках спинного мозга, в вегетативных узлах), норадреналин (в окончаниях симпатических нервных волокон, в гипоталамусе), некоторые аминокислоты и др. Приходящие в окончания аксона нервные импульсы вызывают опорожнение синаптических пузырьков и выведение медиатора в синаптическую щель.
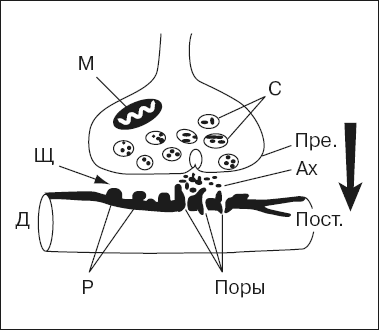
Рис. 2. Схема синапса: Пре. – пресинаптическая мембрана; Пост. – постсинаптическая мембрана; С – синаптические пузырьки; Щ – синаптическая щель; М – митохондрий; Ах – ацетилхолин; Р – рецепторы и поры (Поры) дендрита (Д) следующего нейрона; стрелка – одностороннее проведение возбуждения
По характеру воздействия на последующую нервную клетку различают возбуждающие и тормозящие синапсы.
В возбуждающих синапсах медиаторы (например, ацетилхолин) связываются со специфическими макромолекулами постсинаптической мембраны и вызывают ее деполяризацию. При этом регистрируется небольшое и кратковременное (около 1 мс) колебание мембранного потенциала в сторону деполяризации или возбуждающий постсинаптический потенциал (ВПСП). Для возбуждения нейрона необходимо, чтобы ВПСП достиг порогового уровня. Для этого величина деполяризационного сдвига мембранного потенциала должна составлять не менее 10 мВ. Действие медиатора очень кратковременно (1–2 мс), после чего он расщепляется на неэффективные компоненты (например, ацетилхолин расщепляется ферментом холинэстеразой на холин и уксусную кислоту) или поглощается обратно пресинаптическими окончаниями (например, норадреналин).
В тормозящих синапсах содержатся тормозные медиаторы (например, гамма-аминомасляная кислота). Их действие на постсинаптическую мембрану вызывает усиление выхода ионов калия из клетки и увеличение поляризации мембраны. При этом регистрируется кратковременное колебание мембранного потенциала в сторону гиперполяризации – тормозящий постсинаптический потенциал (ТПСП). В результате нервная клетка оказывается заторможенной. Возбудить ее труднее, чем в исходном состоянии. Для этого понадобится более сильное раздражение, чтобы достичь критического уровня деполяризации.
3.2.4. Возникновение импульсного ответа нейрона
На мембране тела и дендритов нервной клетки находятся как возбуждающие, так и тормозящие синапсы. В отдельные моменты времени часть их может быть неактивной, а другая часть оказывает активное влияние на прилегающие к ним участки мембраны. Общее изменение мембранного потенциала нейрона является результатом сложного взаимодействия (интеграции) местных ВПСП и ТПСП всех многочисленных активированных синапсов. При одновременном влиянии как возбуждающих, так и тормозящих синапсов происходит алгебраическое суммирование (т. е. взаимное вычитание) их эффектов. При этом возбуждение нейрона возникнет лишь в том случае, если сумма возбуждающих постсинаптических потенциалов окажется больше суммы тормозящих. Это превышение должно составлять определенную пороговую величину (около 10 мВ). Только в этом случае появляется потенциал действия клетки. Следует отметить, что в целом возбудимость нейрона зависит от его размеров: чем меньше клетка, тем выше ее возбудимость.
С появлением потенциала действия начинается процесс проведения нервного импульса по аксону и передача его на следующий нейрон или рабочий орган, т. е. осуществляется эффекторная функция нейрона. Нервный импульс является основным средством связи между нейронами.
Таким образом, передача информации в нервной системе происходит с помощью двух механизмов – электрического (ВПСП; ТПСП; потенциал действия) и химического (медиаторы).
3.3. Особенности деятельности нервных центров
Свойства нервных центров в значительной мере связаны с особенностями проведения нервных импульсов через синапсы, связывающие различные нервные клетки.
3.3.1. Особенности проведения возбуждения через нервные центры
Нервным центром называют совокупность нервных клеток, необходимых для осуществления какой-либо функции. Эти центры отвечают соответствующими рефлекторными реакциями на внешнее раздражение, поступившее от связанных с ними рецепторов. Клетки нервных центров реагируют и на непосредственное их раздражение веществами, находящимися в протекающей через них крови (гуморальные влияния). В целостном организме имеется строгое согласование – координация их деятельности.
Проведение волны возбуждения от одного нейрона к другому через синапс происходит в большинстве нервных клеток химическим путем – с помощью медиатора, а медиатор содержится лишь в пресинаптической части синапса и отсутствует в постсинаптической мембране. Поэтому важной особенностью проведения возбуждения через синаптические контакты является одностороннее проведение нервных влияний, которое возможно лишь от пресинаптической мембраны к постсинаптической и невозможно в обратном направлении. В связи с этим поток нервных импульсов в рефлекторной дуге имеет определенное направление от афферентных нейронов к вставочным и затем к эфферентным – мотонейронам или вегетативным нейронам.
Большое значение в деятельности нервной системы имеет другая особенность проведения возбуждения через синапсы – замедленное проведение. Затрата времени на процессы, происходящие от момента подхода нервного импульса к пресинаптической мембране до появления в постсинаптической мембране потенциалов, называется синаптической задержкой. В большинстве центральных нейронов она составляет около 0,3 мс. После этого требуется еще время на развитие возбуждающего постсинаптического потенциала (ВПСП) и потенциала действия. Весь процесс передачи нервного импульса (от потенциала действия одной клетки до потенциала действия следующей клетки) через один синапс занимает примерно 1,5 мс. При утомлении, охлаждении и ряде других воздействий длительность синаптической задержки возрастает. Если же для осуществления какой-либо реакции требуется участие большого числа нейронов (многих сотен и даже тысяч), то суммарная величина задержки проведения по нервным центрам может составить десятые доли секунды и даже целые секунды.

Рис. 3. Схема измерения времени двигательной реакции: А – афферентные, Э – эфферентные и Ц – центральные пути; С – отметка светового сигнала; О – отметка нажима кнопки; t 150 мс – время реакции
При рефлекторной деятельности общее время от момента нанесения внешнего раздражения до появления ответной реакции организма – так называемое скрытое, или латентное, время рефлекса определяется в основном длительностью проведения через синапсы. Величина латентного времени рефлекса служит важным показателем функционального состояния нервных центров. Измерение латентного времени простой двигательной реакции человека на внешний сигнал широко используется в практике для оценки функционального состояния ЦНС (рис. 3).
3.3.2. Суммация возбуждения
В ответ на одиночную афферентную волну, идущую от рецепторов к нейронам, в пресинаптической части синапса освобождается небольшое количество медиатора. При этом в постсинаптической мембране нейрона обычно возникает ВПСП – небольшая местная деполяризация. Для того чтобы общая по всей мембране нейрона величина ВПСП достигала порога возникновения потенциала действия, требуется суммация на мембране клетки многих подпороговых ВПСП. Лишь в результате такой суммации возбуждения возникает ответ нейрона. Различают пространственную и временную суммацию.
Пространственная суммация наблюдается в случае одновременного поступления нескольких импульсов в один и тот же нейрон по разным пресинаптическим волокнам. Одномоментное возбуждение синапсов в различных участках мембраны нейрона повышает амплитуду суммарного ВПСП до пороговой величины. В результате возникает ответный импульс нейрона и осуществляется рефлекторная реакция. Например, для получения ответа двигательной клетки спинного мозга обычно требуется одновременная активация 50–100 афферентных волокон от соответствующих периферических рецепторов.
Временная суммация происходит при активации одного и того же афферентного пути серией последовательных раздражений. Если интервалы между поступающими импульсами достаточно коротки и ВПСП нейрона от предыдущих раздражений не успевают затухать, то последующие ВПСП накладываются друг на друга, пока деполяризация мембраны нейрона не достигнет критического уровня для возникновения потенциала действия. Таким способом даже слабые раздражения через некоторое время могут вызывать ответные реакции организма (например, чихание и кашель в ответ на слабые раздражения слизистой оболочки дыхательных путей).
3.3.3. Трансформация и усвоение ритма
Характер ответного разряда нейрона зависит не только от свойств раздражителя, но и от функционального состояния самого нейрона (его мембранного заряда, возбудимости, лабильности). Нервные клетки обладают свойством изменять частоту передающихся импульсов, т. е. свойством трансформации ритма.
При высокой возбудимости нейрона (например, после приема кофеина) может возникать учащение импульсации (мультипликация ритма), а при низкой возбудимости (например, при утомлении) происходит урежение ритма, так как несколько приходящих импульсов должны суммироваться, чтобы наконец достичь порога возникновения потенциала действия. Эти изменения частоты импульсации могут усиливать или ослаблять ответные реакции организма на внешние раздражения.
При ритмических раздражениях активность нейрона может настроиться на ритм приходящих импульсов, т. е. наблюдается явление усвоения ритма (Ухтомский А. А., 1928). Развитие усвоения ритма обеспечивает сонастройку активности многих нервных центров при управлении сложными двигательными актами, особенно это важно для поддержания темпа циклических упражнений.
3.3.4. Следовые процессы
После окончания действия раздражителя активное состояние нервной клетки или нервного центра обычно продолжается еще некоторое время. Длительность следовых процессов различна: небольшая в спинном мозге (несколько секунд или минут), значительно больше в центрах головного мозга (десятки минут, часы или даже дни) и очень большая в коре больших полушарий (до нескольких десятков лет).
Поддерживать явное и кратковременное состояние возбуждения в нервном центре могут импульсы, циркулирующие по замкнутым цепям нейронов. Значительно сложнее по природе длительно сохраняющиеся скрытые следы. Предполагают, что длительное сохранение в нервной клетке следов со всеми характерными свойствами раздражителя основано на изменении структуры составляющих клетку белков и на перестройке синаптических контактов.
Непродолжительные импульсные последействия (длительностью до 1 часа) лежат в основе так называемой кратковременной памяти, а длительные следы, связанные со структурными и биохимическими перестройками в клетках, – в основе формирования долговременной памяти.
3.4. Координация деятельности ЦНС
Процессы координации деятельности ЦНС основаны на согласовании двух нервных процессов – возбуждения и торможения. Торможение является активным нервным процессом, который предупреждает или угнетает возбуждение.
3.4.1. Значение процесса торможения в ЦНС
Явление торможения в нервных центрах было впервые открыто И. М. Сеченовым в 1862 г. Значение этого процесса было рассмотрено им в книге «Рефлексы головного мозга» (1863).
Опуская лапку лягушки в кислоту и одновременно раздражая некоторые участки головного мозга (например, накладывая кристаллик поваренной соли на область промежуточного мозга), И. М. Сеченов наблюдал резкую задержку и даже полное отсутствие «кислотного» рефлекса спинного мозга (отдергивания лапки). Отсюда он сделал заключение, что одни нервные центры могут существенно изменять рефлекторную деятельность в других центрах, в частности вышележащие нервные центры могут тормозить деятельность нижележащих. Описанный опыт вошел в историю физиологии под названием Сеченовское торможение.
Тормозные процессы – необходимый компонент в координации нервной деятельности. Во-первых, процесс торможения ограничивает распространение возбуждения на соседние нервные центры, чем способствует его концентрации в необходимых участках нервной системы. Во-вторых, возникая в одних нервных центрах параллельно с возбуждением других нервных центров, процесс торможения тем самым выключает деятельность ненужных в данный момент органов. В-третьих, развитие торможения в нервных центрах предохраняет их от чрезмерного перенапряжения при работе, т. е. играет охранительную роль.
3.4.2. Постсинаптическое и пресинаптическое торможение
Процесс торможения, в отличие от возбуждения, не может распространяться по нервному волокну – это всегда местный процесс в области синаптических контактов. По месту возникновения различают пресинаптическое и постсинаптическое торможение.
Постсинаптическое торможение – это тормозные эффекты, возникающие в постсинаптической мембране. Чаще всего этот вид торможения связан с наличием в ЦНС специальных тормозных нейронов. Они представляют собой особый тип вставочных нейронов, у которых окончания аксонов выделяют тормозный медиатор. Одним из таких медиаторов является гамма-аминомасляная кислота (ГАМК).
Нервные импульсы, подходя к тормозным нейронам, вызывают в них такой же процесс возбуждения, как и в других нервных клетках. В ответ по аксону тормозной клетки распространяется обычный потенциал действия. Однако, в отличие от других нейронов, окончания аксона при этом выделяют не возбуждающий, а тормозной медиатор. В результате тормозные клетки тормозят те нейроны, на которых оканчиваются их аксоны.
К специальным тормозным нейронам относятся клетки Рэншоу в спинном мозге, клетки Пуркинье мозжечка, корзинчатые клетки в промежуточном мозге и др. Большое значение, например, тормозные клетки имеют при регуляции деятельности мышц-антагонистов: приводя к расслаблению мышц-антагонистов, они облегчают тем самым одновременное сокращение мышц-агонистов (рис. 4).
Клетки Рэншоу участвуют в регуляции уровня активности отдельных мотонейронов спинного мозга. При возбуждении мотонейрона импульсы поступают по его аксону к мышечным волокнам и одновременно по коллатералям аксона – к тормозной клетке Рэншоу. Аксоны последней «возвращаются» к этому же нейрону, вызывая его торможение. Чем больше возбуждающих импульсов посылает мотонейрон на периферию (а значит, и к тормозной клетке), тем сильнее это возвратное торможение (разновидность постсинаптического торможения). Такая замкнутая система действует как механизм саморегуляции нейрона, предохраняя его от чрезмерной активности.
Клетки Пуркинье мозжечка своими тормозными влияниями на клетки подкорковых ядер и стволовых структур участвуют в регуляции тонуса мышц.

Рис. 4. Участие тормозной клетки в регуляции мышц-антагонистов: В и Т – возбуждающий и тормозной нейроны; возбуждение (+) мотонейрона мышцы-сгибателя (МС) и торможение (-) мотонейрона мышцы-разгибателя (MP); Р – кожный рецептор
Корзинчатые клетки в промежуточном мозге являются как бы воротами, которые пропускают или не пропускают импульсы, идущие в кору больших полушарий от различных областей тела.
Пресинаптическое торможение возникает перед синаптическим контактом – в пресинаптической области. Окончание аксона тормозной нервной клетки образует синапс на конце аксона возбуждающей нервной клетки, вызывает чрезмерно сильную деполяризацию мембраны этого аксона, которая угнетает проходящие здесь потенциалы действия и тем самым блокирует передачу возбуждения. Этот вид торможения ограничивает поток афферентных импульсов к нервным центрам, выключая посторонние для основной деятельности влияния.
3.4.3. Явления иррадиации и концентрации
При раздражении одного рецептора возбуждение может в принципе распространяться в ЦНС в любом направлении и на любую нервную клетку. Это происходит благодаря многочисленным взаимосвязям нейронов одной рефлекторной дуги с нейронами других рефлекторных дуг. Распространение процесса возбуждения на другие нервные центры называют явлением иррадиации.
Чем сильнее афферентное раздражение и чем выше возбудимость окружающих нейронов, тем больше нейронов охватывает процесс иррадиации. Процессы торможения ограничивают иррадиацию и способствуют концентрации возбуждения в исходном пункте ЦНС.
Процесс иррадиации играет важную положительную роль при формировании новых реакций организма (ориентировочных реакций, условных рефлексов). Чем больше активируется различных нервных центров, тем легче отобрать из их числа наиболее нужные для последующей деятельности центры. Благодаря иррадиации возбуждения между различными нервными центрами возникают новые функциональные взаимосвязи – условные рефлексы. На этой основе возможно, например, формирование новых двигательных навыков.
Вместе с тем иррадиация возбуждения может оказать и отрицательное воздействие на состояние и поведение организма, нарушая тонкие взаимоотношения между возбужденными и заторможенными нервными центрами и вызывая нарушения координации движений.
3.4.4. Доминанта
Исследуя особенности межцентральных отношений, А. А. Ухтомский обнаружил, что если в организме животного осуществляется сложная рефлекторная реакция, например повторяющиеся акты глотания, то электрическое раздражение моторных центров не только перестает вызывать в этот момент движение конечностей, но и усиливает протекание начавшейся цепной реакции глотания, которая оказалась главенствующей.
Такой господствующий очаг возбуждения в ЦНС, определяющий текущую деятельность организма, А. А. Ухтомский (1923) обозначил термином доминанта.
Доминирующий очаг может возникнуть при повышенном уровне возбудимости нервных клеток, который создается различными гуморальными и нервными влияниями. Он подавляет деятельность других центров, оказывая сопряженное торможение.
Объединение большого числа нейронов в одну доминантную систему происходит путем взаимного сонастраивания на общий темп активности, т. е. путем усвоения ритма. Одни нервные клетки снижают свой более высокий темп деятельности, а другие – повышают низкий темп до некоторого среднего, оптимального ритма. Доминанта может надолго сохраняться в скрытом, следовом состоянии (потенциальная доминанта). При возобновлении прежнего состояния или прежней внешней ситуации доминанта может снова возникнуть (актуализация доминанты). Например, в предстартовом состоянии активизируются все те нервные центры, которые входили в рабочую систему во время предыдущих тренировок, и соответственно усиливаются функции, связанные с работой. Мысленное выполнение физических упражнений или представление движений также воспроизводит рабочую доминанту, что обеспечивает тренирующий эффект представления движений и является основой так называемой идеомоторной тренировки. При полном расслаблении (например, при аутогенной тренировке) спортсмены добиваются устранения рабочих доминант, что ускоряет процессы восстановления.
Как фактор поведения, доминанта связана с высшей нервной деятельностью и психологией человека. Доминанта является физиологической основой акта внимания. При наличии доминанты многие влияния внешней среды остаются вне нашего внимания, но зато более интенсивно улавливаются и анализируются те, которые нас особенно интересуют. Таким образом, доминанта является мощным фактором отбора биологически и социально наиболее значимых раздражений.
3.5. Функции спинного мозга и подкорковых отделов головного мозга
В ЦНС различают более древние сегментарные и эволюционно более молодые надсегментарные отделы нервной системы. К сегментарным отделам относят спинной, продолговатый и средний мозг, участки которых регулируют функции отдельных частей тела, лежащих на том же уровне. Надсегментарные отделы – промежуточный мозг, мозжечок и кора больших полушарий не имеют непосредственных связей с органами тела, а управляют их деятельностью через нижележащие сегментарные отделы.
3.5.1. Спинной мозг
Спинной мозг является низшим и наиболее древним отделом ЦНС.
В составе серого вещества спинного мозга человека насчитывают около 13,5 млн нервных клеток. Из них основную массу (97 %) представляют промежуточные клетки (вставочные или интернейроны), которые обеспечивают сложные процессы координации внутри спинного мозга. Среди мотонейронов спинного мозга выделяют крупные альфа-мотонейроны и мелкие – гамма-мотонейроны. От альфа-мотонейронов отходят наиболее толстые и быстропроводящие волокна двигательных нервов, вызывающие сокращения скелетных мышечных волокон. Тонкие волокна гамма-мотонейронов не вызывают сокращения мышц. Они подходят к проприорецепторам – мышечным веретенам – и регулируют их чувствительность.
Рефлексы спинного мозга можно подразделить на двигательные, осуществляемые альфа-мотонейронами передних рогов, и вегетативные, осуществляемые афферентными клетками боковых рогов.
Мотонейроны спинного мозга иннервируют все скелетные мышцы (за исключением мышц лица). Спинной мозг осуществляет элементарные двигательные рефлексы – сгибательные и разгибательные, ритмические, шагательные, возникающие при раздражении кожи или проприорецепторов мышц и сухожилий, а также посылает постоянную импульсацию к мышцам, поддерживая мышечный тонус. Специальные мотонейроны иннервируют дыхательную мускулатуру (межреберные мышцы и диафрагму) и обеспечивают дыхательные движения. Вегетативные нейроны иннервируют все внутренние органы (сердце, сосуды, потовые железы, железы внутренней секреции, пищеварительный тракт, мочеполовую систему).
Проводниковая функция спинного мозга связана с передачей в вышележащие отделы нервной системы получаемого с периферии потока информации и с проведением импульсов, идущих из головного мозга в спинной.
За последние годы разработаны специальные методики для изучения деятельности спинного мозга у здорового человека. Так, например, функциональное состояние альфа-мотонейронов оценивают по изменению ответных потенциалов мышц при периферических раздражениях – так называемому Н-рефлексу (рефлексу Гофмана) икроножной мышцы при раздражении большеберцового нерва и по Т-рефлексу (от тендон – «сухожилие») камбаловидной мышцы при раздражении ахиллова сухожилия. Разработаны методики регистрации (с неповрежденных покровов тела) потенциалов, проходящих по спинному мозгу в головной.
3.5.2. Продолговатый мозг и варолиев мост
Продолговатый мозг и варолиев мост (в целом – задний мозг) являются частью ствола мозга. Здесь находится большая группа черепномозговых нервов (от V до XII пары), иннервирующих кожу, слизистые оболочки, мускулатуру головы и ряд внутренних органов (сердце, легкие, печень). Тут же находятся центры многих пищеварительных рефлексов – жевания, глотания, движений желудка и части кишечника, выделения пищеварительных соков, а также центры некоторых защитных рефлексов (чихания, кашля, мигания, слезоотделения, рвоты) и центры водно-солевого и сахарного обмена. На дне IV желудочка в продолговатом мозге находится жизненно важный дыхательный центр, состоящий из центров вдоха и выдоха. Его составляют мелкие клетки, посылающие импульсы к дыхательным мышцам через мотонейроны спинного мозга.
В непосредственной близости расположен сердечно-сосудистый центр. Его крупные клетки регулируют деятельность сердца и просвет сосудов. Переплетение клеток дыхательного и сердечно-сосудистого центров обеспечивает их тесное взаимодействие.
Продолговатый мозг играет важную роль в осуществлении двигательных актов и в регуляции тонуса скелетных мышц, повышая тонус мышц-разгибателей. Он принимает участие, в частности, в осуществлении установочных рефлексов позы (шейных, лабиринтных). Через продолговатый мозг проходят восходящие пути слуховой, вестибулярной, проприоцептивной и тактильной чувствительности.
3.5.3. Средний мозг
В состав среднего мозга всходят четверохолмия, черная субстанция и красные ядра. В передних буграх четверохолмия находятся зрительные подкорковые центры, а в задних – слуховые. Средний мозг участвует в регуляции движений глаз, осуществляет зрачковый рефлекс (расширение зрачков в темноте и сужение их на свету).
Четверохолмия выполняют ряд реакций, являющихся компонентами ориентировочного рефлекса. В ответ на внезапное раздражение происходит поворот головы и глаз в сторону раздражителя, а у животных – настораживание ушей. Этот рефлекс (по И. П. Павлову, рефлекс «Что такое?») необходим для подготовки организма к своевременной реакции на любое новое воздействие.
Черная субстанция среднего мозга имеет отношение к рефлексам жевания и глотания, участвует в регуляции тонуса мышц (особенно при выполнении мелких движений пальцами рук) и в организации содружественных двигательных реакций.
Красное ядро среднего мозга выполняет моторные функции – регулирует тонус скелетных мышц, вызывая усиление тонуса мышц-сгибателей. Оказывая значительное влияние на тонус скелетных мышц, средний мозг принимает участие в ряде установочных рефлексов поддержания позы (выпрямительных – установке тела теменем вверх и др.).
3.5.4. Промежуточный мозг
В состав промежуточного мозга входят таламус (зрительные бугры) и гипоталамус (подбугорье).
Через таламус проходят все афферентные пути (за исключением обонятельных), которые направляются в соответствующие воспринимающие области коры (слуховые, зрительные и пр.). Ядра таламуса подразделяются на специфические и неспецифические. К специфическим относят переключательные (релейные) ядра и ассоциативные. Через переключательные ядра таламуса передаются афферентные влияния от всех рецепторов тела. Ассоциативные ядра получают импульсы от переключательных ядер и обеспечивают их взаимодействие. Помимо этих ядер в таламусе имеются неспецифические ядра, которые оказывают как активирующие, так и тормозящие влияния на небольшие области коры.
Благодаря обширным связям таламус играет важнейшую роль в жизнедеятельности организма. Импульсы, идущие от таламуса в кору, изменяют состояние корковых нейронов и регулируют ритм корковой активности. С непосредственным участием таламуса происходит образование условных рефлексов и выработка двигательных навыков, формирование эмоций человека, его мимики. Таламусу принадлежит большая роль в возникновении ощущений, в частности ощущения боли. С его деятельностью связывают регуляцию биоритмов в жизни человека (суточных, сезонных и др.).
Гипоталамус является высшим подкорковым центром регуляции вегетативных функций, состояний бодрствования и сна. Здесь расположены вегетативные центры, регулирующие обмен веществ в организме, обеспечивающие поддержание постоянства температуры тела (у теплокровных) и нормального уровня кровяного давления, поддерживающие водный баланс, регулирующие чувство голода и насыщения. Раздражения задних ядер гипоталамуса вызывает усиление симпатических влияний, а передних – парасимпатические эффекты.
Благодаря связи гипоталамуса с гипофизом (гипоталамогипофизарная система) осуществляется контроль деятельности желез внутренней секреции. Вегетативные и гормональные реакции, регулируемые гипоталамусом, являются компонентами эмоциональных и двигательных реакций человека.
3.5.5. Неспецифическая система мозга
Неспецифическая система занимает срединную часть ствола мозга. Она не связана с анализом какой-либо специфической чувствительности или с выполнением определенных рефлекторных реакций. Импульсы в эту систему поступают через боковые ответвления от всех специфических путей, в результате чего обеспечивается их обширное взаимодействие. Для неспецифической системы характерно расположение нейронов в виде диффузной сети, обилие и разнообразие их отростков. В связи с этим она и получила название сетевидного образования, или ретикулярной формации.
Различают два типа влияния неспецифической системы на работу других нервных центров – активирующее и тормозящее. Оба типа этих влияний могут быть восходящими (к вышележащим центрам) и нисходящими (к нижележащим центрам). Они служат для регулирования функционального состояния мозга, уровня бодрствования и регуляции позно-тонических и фазных реакций скелетных мышц.
3.5.6. Мозжечок
Мозжечок – надсегментарное образование, не имеющее непосредственных связей с исполнительными аппаратами. Мозжечок состоит из непарного образования – червя и парных полушарий.
Основными нейронами коры мозжечка являются многочисленные клетки Пуркинье. Благодаря обширным связям (на каждой клетке оканчивается до 200 000 синапсов) в них происходит интеграция самых различных сенсорных влияний, в первую очередь проприоцептивных, тактильных и вестибулярных. Представительство разных периферических рецепторов в коре мозжечка имеет соматотопическую организацию (греч. соматос – «тело», топос – «место»), т. е. отражает порядок их расположения в теле человека. Кроме того, этот порядок расположения соответствует такому же порядку расположения представительства участков тела в коре больших полушарий, что облегчает обмен информацией между корой и мозжечком и обеспечивает их совместную деятельность в управлении поведением человека. Правильная геометрическая организация нейронов мозжечка обусловливает его значение в отсчете времени и четком поддержании темпа циклических движений.
Основной функцией мозжечка является регуляция позно-тонических реакций и координация двигательной деятельности (Орбели Л. А., 1926).
По анатомическим особенностям (связям коры мозжечка с его ядрами) и функциональному значению мозжечок подразделяют на три продольные зоны:
• внутреннюю, или медиальную, – кору червя, функцией которой является регуляция тонуса скелетных мышц, поддержание позы и равновесия тела;
• промежуточную – среднюю часть коры полушарий мозжечка, функция которой состоит в согласовании позных реакций с движениями и коррекции ошибок;
• боковую, или латеральную, кору полушарий мозжечка, которая совместно с промежуточным мозгом и корой больших полушарий участвует в программировании быстрых баллистических движений (бросков, ударов, прыжков и пр.).
3.5.7. Базальные ядра
К базальным ядрам относят полосатое тело, состоящее из хвостатого ядра и скорлупы, и бледное ядро, а в настоящее время причисляют также миндалевидное тело (относящееся к вегетативным центрам лимбической системы) и черную субстанцию среднего мозга.
Афферентные влияния приходят к базальным ядрам от рецепторов тела через таламус и от всех областей коры больших полушарий. Они почти исключительно поступают в полосатое тело. Эфферентные влияния от него направляются к бледному ядру и далее к стволовым центрам экстрапирамидной системы, а также через таламус обратно к коре.
Базальные ядра участвуют в образовании условных рефлексов и осуществлении сложных безусловных рефлексов (оборонительных, пищедобывательных и др.). Они обеспечивают необходимое положение тела во время физической работы, а также протекание автоматических ритмических движений (древних автоматизмов).
Бледное ядро выполняет основную моторную функцию, а полосатое тело регулирует его активность. В настоящее время выявлено значение хвостатого ядра в контроле сложных психических процессов – внимания, памяти, обнаружении ошибок.
3.6. Вегетативная нервная система
Все функции организма условно можно разделить на соматические, или анимальные (животные), связанные с восприятием внешней информации и деятельностью мышц, и вегетативные (растительные), связанные с деятельностью внутренних органов, – процессы дыхания, кровообращения, пищеварения, выделения, обмена веществ, роста и размножения.
3.6.1. Функциональная организация вегетативной нервной системы
Вегетативной нервной системой называют совокупность эфферентных нервных клеток спинного и головного мозга, а также клеток особых узлов (ганглиев), иннервирующих внутренние органы. Раздражения различных рецепторов тела могут вызвать изменения соматических и вегетативных функций, так как афферентные и центральные отделы этих рефлекторных дуг общие. Они различаются лишь своими эфферентными отделами. Характерной особенностью эфферентных путей, входящих в рефлекторные дуги вегетативных рефлексов, является их двухнейронное строение (один нейрон находится в ЦНС, другой – в ганглиях или в иннервируемом органе).
Вегетативная нервная система подразделяется на два отдела – симпатический и парасимпатический (рис. 5).
Эфферентные пути симпатической нервной системы начинаются в грудном и поясничном отделах спинного мозга от нейронов его боковых рогов. Передача возбуждения с предузловых симпатических волокон на послеузловые происходит с участием медиатора ацетилхолина, а с послеузловых волокон на иннервируемые органы – с участием медиатора норадреналина. Исключением являются волокна, иннервирующие потовые железы и расширяющие сосуды скелетных мышц, где возбуждение передается с помощью ацетилхолина.

Рис. 5. Вегетативная нервная система. Слева – область выхода волокон: парасимпатической (черный цвет) и симпатической (заштриховано) систем. Арабские цифры – номера грудных сегментов, римские – номера поясничных сегментов. Справа – строение эфферентной части рефлекторной дуги вегетативных рефлексов
Эфферентные пути парасимпатической нервной системы начинаются в головном мозге (от некоторых ядер среднего и продолговатого мозга) и в спинном мозге (от нейронов крестцового отдела). Проведение возбуждения в синапсах парасимпатического пути происходит с участием медиатора ацетилхолина. Второй эфферентный нейрон находится в иннервируемом органе или вблизи от него.
Высшим регулятором вегетативных функций является гипоталамус, который действует совместно с ретикулярной формацией и лимбической системой под контролем коры больших полушарий. Кроме того, нейроны, расположенные в самих органах или в симпатических узлах, могут осуществлять собственные рефлекторные реакции без участия ЦНС – «периферические рефлексы».
3.6.2. Функции симпатической нервной системы
С участием симпатической нервной системы протекают многие важные рефлексы в организме, направленные на обеспечение его деятельного состояния, в том числе – его двигательной активности. К ним относятся рефлексы расширения бронхов, учащения и усиления сердечных сокращений, расширения сосудов сердца и легких при одновременном сужении сосудов кожи и органов брюшной полости (обеспечение перераспределения крови), выброс депонированной крови из печени и селезенки, расщепление гликогена до глюкозы в печени (мобилизация углеводных источников энергии), усиление деятельности желез внутренней секреции и потовых желез. Симпатическая нервная система снижает деятельность ряда внутренних органов: в результате сужения сосудов в почках уменьшаются процессы мочеобразования, угнетается секреторная и моторная деятельность органов желудочно-кишечного тракта; предотвращается акт мочеиспускания – расслабляется мышца стенки мочевого пузыря и сокращается его сфинктер.
Повышенная активность организма сопровождается симпатическим рефлексом расширения зрачка. Огромное значение для двигательной деятельности организма имеет трофическое влияние симпатических нервов на скелетные мышцы, улучшающее их обмен веществ и функциональное состояние, снимающее утомление.
Симпатический отдел нервной системы не только повышает уровень функционирования организма, но и мобилизует его скрытые функциональные резервы, активирует деятельность мозга, повышает защитные реакции (иммунные реакции, барьерные механизмы и др.), запускает гормональные реакции. Особенное значение имеет симпатическая нервная система при развитии стрессовых состояний в наиболее сложных условиях жизнедеятельности. Л. А. Орбели подчеркивал важнейшее значение симпатических влияний для приспособления (адаптации) организма к напряженной работе, к различным условиям внешней среды. Эта функция была им названа адаптационно-трофической.
3.6.3. Функции парасимпатической нервной системы
Парасимпатическая нервная система осуществляет сужение бронхов; замедление и ослабление сердечных сокращений; сужение сосудов сердца; пополнение энергоресурсов (синтез гликогена в печени и усиление процессов пищеварения); усиление процессов мочеобразования в почках и обеспечение акта мочеиспускания (сокращение мышц мочевого пузыря и расслабление его сфинктера) и др. Парасимпатическая нервная система преимущественно оказывает пусковые влияния: сужение зрачка, бронхов, включение деятельности пищеварительных желез и т. п.
Деятельность парасимпатического отдела вегетативной нервной системы направлена на текущую регуляцию функционального состояния, на поддержание постоянства внутренней среды – гомеостаза. Парасимпатический отдел обеспечивает восстановление различных физиологических показателей, резко измененных после напряженной мышечной работы, пополнение израсходованных энергоресурсов. Медиатор парасимпатической системы – ацетилхолин, снижая чувствительность адренорецепторов к действию адреналина и норадреналина, оказывает определенное антистрессорное влияние.
3.6.4. Вегетативные рефлексы
Через вегетативные симпатические и парасимпатические пути ЦНС осуществляет некоторые вегетативные рефлексы, начинающиеся с различных рецепторов внешней и внутренней среды: висцеро-висцеральные (с внутренних органов на внутренние органы – например, дыхательно-сердечный рефлекс); дермо-висцеральные (с кожных покровов – изменение деятельности внутренних органов при раздражении активных точек кожи, например, иглоукалыванием, точечным массажем); с рецепторов глазного яблока – глазо-сердечный рефлекс Ашнера (урежение сердцебиений при надавливании на глазные яблоки – парасимпатический эффект); моторно-висцеральные – например, ортостатическая проба (учащение сердцебиения при переходе из положения лежа в положение стоя – симпатический эффект) и др. (рис. 6). Они используются для оценки функционального состояния организма и особенно состояния вегетативной нервной системы (оценки влияния симпатического или парасимпатического ее отдела).

Рис. 6. Вегетативные рефлексы. Влияние положения тела на частоту сердечных сокращений (уд./мин) (по: М. Р. Могендович, 1972)
3.7. Лимбическая система
Под лимбической системой понимают ряд корковых и подкорковых структур, функции которых связаны с организацией мотивационно-эмоциональных реакций, процессами памяти и обучения.
Корковые отделы лимбической системы, представляющие ее высший отдел, находятся на нижних и внутренних поверхностях больших полушарий (участки лобной коры, поясная извилина или лимбическая кора, гиппокамп и др.). К подкорковым структурам лимбической системы относят гипоталамус, некоторые ядра таламуса, среднего мозга и ретикулярной формации. Между всеми этими отделами имеются тесные прямые и обратные связи, образующие так называемое «лимбическое кольцо».
Лимбическая система участвует в самых разнообразных проявлениях деятельности организма – в регуляции пищевого и питьевого поведения, цикла сон – бодрствование, в процессах формирования памятного следа (отложения и извлечения из памяти), в развитии агрессивно-оборонительных реакций, обеспечивая избирательный характер поведения. Она формирует положительные и отрицательные эмоции со всеми двигательными, вегетативными и гормональными их компонентами. Электрические раздражения различных участков лимбической системы через вживленные электроды (в экспериментах на животных, в клинике при лечении больных) выявили наличие центров удовольствия, формирующих положительные эмоции, и неудовольствия, формирующих отрицательные эмоции. Изолированное раздражение таких точек в глубоких структурах мозга человека вызывало появление чувства «беспричинной радости», «беспредметной тоски», «безотчетного страха».
3.8. Функции коры больших полушарий
У высших млекопитающих животных и человека ведущий отдел ЦНС – это кора больших полушарий.
3.8.1. Корковые нейроны
Кора представляет собой слой серого вещества толщиной 2–3 мм, содержащий в среднем около 14 млрд нервных клеток. Характерным в ней является обилие межнейронных связей, рост которых продолжается до 18 лет, а в ряде случаев и далее.
Основными типами корковых клеток являются пирамидные и звездчатые нейроны. Звездчатые нейроны связаны с процессами восприятия раздражений и объединением деятельности различных пирамидных нейронов.
Пирамидные нейроны осуществляют эфферентную функцию коры (преимущественно через пирамидный тракт) и внутрикорковые процессы взаимодействия между удаленными друг от друга нейронами. Наиболее крупные пирамидные клетки – гигантские пирамиды Беца – находятся в передней центральной извилине (моторной зоне коры).
Функциональной единицей коры является вертикальная колонка взаимосвязанных нейронов. Вытянутые по вертикали крупные пирамидные клетки с расположенными над ними и под ними нейронами образуют функциональные объединения нейронов. Все нейроны вертикальной колонки отвечают на одно и то же афферентное раздражение (от одного и того же рецептора) одинаковой реакцией и совместно формируют эфферентные ответы пирамидных нейронов. По мере надобности вертикальные колонки могут объединяться в более крупные образования, обеспечивая сложные реакции.
3.8.2. Функциональное значение различных корковых полей
По особенностям строения и функциональному значению отдельных корковых участков вся кора подразделяется на три основные группы полей – первичные, вторичные и третичные (рис. 7).
Первичные поля связаны с органами чувств и органами движения на периферии. Они обеспечивают возникновение ощущений. К ним относятся, например, поле болевой и мышечно-суставной чувствительности в задней центральной извилине коры, зрительное поле в затылочной области, слуховое поле в височной области и моторное поле в передней центральной извилине. В первичных полях находятся высокоспециализированные клетки-определители, или детекторы, избирательно реагирующие только на определенные раздражения. Например, в зрительной коре имеются нейроны-детекторы, возбуждающиеся только при включении или при выключении света, чувствительные лишь к определенной его интенсивности, к конкретным интервалам светового воздействия, к определенной длине волны и т. д. При разрушении первичных полей коры возникают так называемые корковая слепота, корковая глухота и т. п.

Рис. 7. Первичные, вторичные и третичные поля коры больших полушарий. А: крупные точки – первичные поля, средние – вторичные поля мелкие – третичные поля; Б: первичные (проекционные) поля коры больших полушарий
Вторичные поля расположены рядом с первичными. В них происходит осмысливание и узнавание звуковых, световых и других сигналов, возникают сложные формы обобщенного восприятия. При поражении вторичных полей сохраняется способность видеть предметы, слышать звуки, но человек их не узнает, не помнит значения.
Третичные поля развиты практически только у человека. Это ассоциативные области коры, обеспечивающие высшие формы анализа и синтеза и формирующие целенаправленную поведенческую деятельность человека. Третичные поля находятся: в задней половине коры – между теменными, затылочными и височными областями; в передней половине – в передних частях лобных областей. Их роль особенно велика в организации согласованной работы обоих полушарий. Третичные поля созревают у человека позже других корковых полей и раньше других деградируют при старении.
Функцией задних третичных полей (главным образом, нижнетеменных областей коры) является прием, переработка и хранение информации. Они формируют представление о схеме тела и схеме пространства, обеспечивая пространственную ориентацию движений. Передние третичные поля (переднелобные области) выполняют общую регуляцию сложных форм поведения человека, формируя намерения и планы, программы произвольных движений и контроль за их выполнением. Развитие третичных полей у человека связывают с функцией речи. Мышление (внутренняя речь) возможно только при совместной деятельности различных сенсорных систем, объединение информации от которых происходит в третичных полях. При врожденном недоразвитии третичных полей человек не в состоянии овладеть речью (произносит лишь бессмысленные звуки) и даже простейшими двигательными навыками (не может одеваться, пользоваться орудиями труда и т. п.).
3.8.3. Парная деятельность и доминирование полушарий
Обработка информации осуществляется в результате парной деятельности обоих полушарий головного мозга. Однако, как правило, одно из полушарий является ведущим – доминантным. У большинства людей с ведущей правой рукой (правшей) доминантным является левое полушарие, а соподчиненным (субдоминантным) – правое полушарие.
Левое полушарие по сравнению с правым имеет более тонкое нейронное строение, большее богатство взаимосвязей нейронов, более концентрированное представительство функций и лучшие условия кровоснабжения. В левом доминантном полушарии находится моторный центр речи (центр Брока), обеспечивающий речевую деятельность, и сенсорный центр речи, осуществляющий понимание слов. Левое полушарие специализировано на тонком сенсомоторном контроле за движениями рук.
У человека различают три формы функциональной асимметрии: моторную, сенсорную и психическую. Как правило, у человека имеются ведущая рука, нога, глаз и ухо. Однако проблема функциональной асимметрии довольно сложна. Например, у человека-правши может быть ведущим левый глаз или левое ухо, сигналы от которых являются главенствующими. При этом в каждом полушарии могут быть представлены функции не только противоположной, но и одноименной стороны тела. В результате этого обеспечивается возможность замещения одного полушария другим в случае его повреждения, а также создается структурная основа для переменного доминирования полушарий при управлении движениями.
Психическая асимметрия проявляется в виде определенной специализации полушарий. Для левого полушария характерны аналитические процессы, последовательная обработка информации, в том числе с помощью речи, абстрактное мышление, оценка временных отношений, предвосхищение будущих событий, успешное решение вербально-логических задач. В правом полушарии информация обрабатывается целостно, синтетически (без расчленения на детали), с учетом прошлого опыта и без участия речи, преобладает предметное мышление. Эти особенности позволяют связывать с правым полушарием восприятие пространственных признаков и решение зрительно-пространственных задач. Функции правого полушария связаны с прошедшим временем, а левого – с будущим.
3.8.4. Электрическая активность коры больших полушарий
Изменения функционального состояния коры отражаются в записи ее электрической активности – электроэнцефалограммы (ЭЭГ). Современные электроэнцефалографы усиливают потенциалы мозга в 2–3 млн раз и дают возможность исследовать ЭЭГ от многих точек коры одновременно, т. е. изучать системные процессы. Регистрация ЭЭГ производится в виде чернильной записи на бумаге, а также в виде целостной картины на схеме поверхности мозга, т. е. карты мозга (метод картирования) на экране мониторов современных компьютерных систем (рис. 8).

Рис. 8. Картирование мозга: многоканальная регистрация электроэнцефалограммы (ЭЭГ) человека на экране монитора и отражение возбужденных (светлые зоны) и заторможенных (темные зоны) участков коры

Рис. 9. ЭЭГ затылочной (а – д) и моторной (е – з) областей коры больших полушарий человека при различных состояниях и во время мышечной работы: а – активное состояние, глаза открыты (бета-ритм); б – покой, глаза закрыты (альфа-ритм); в – дремота (тета-ритм); г – засыпание (медленные волны); д – глубокий сон (дельта-ритм); е – непривычная или тяжелая работа – асинхронная частая активность (явление десинхронизиции); ж – циклическая работа – медленные потенциалы в темпе движений («меченые ритмы» ЭЭГ); з – выполнение освоенного движения – появление альфа-ритма
Различают определенные диапазоны частот, называемые ритмами ЭЭГ (рис. 9): в состоянии относительного покоя чаще всего регистрируется альфа-ритм (8–13 колебаний в 1 с); в состоянии активного внимания – бета-ритм (14 колебаний в 1 с и выше); при засыпании, некоторых эмоциональных состояниях – тета-ритм (4–7 колебаний в 1 с); при глубоком сне, потере сознания, наркозе – дельта-ритм (1–3 колебания в 1 с).
В ЭЭГ отражаются особенности взаимодействия корковых нейронов при умственной и физической работе (Ливанов М. Н., 1972). Отсутствие налаженной координации при выполнении непривычной или тяжелой работы приводит к так называемой десинхронизации ЭЭГ – быстрой асинхронной активности. По мере формирования двигательного навыка в ЭЭГ возникают явления синхронизации ЭЭГ – усиления взаимосвязанности (синхронности и синфазности) электрической активности различных областей коры, участвующих в управлении движениями. При циклической работе появляются медленные потенциалы в темпе выполняемого, воображаемого или предстоящего движения – «меченые ритмы» (Сологуб Е. Б., 1973).
Помимо фоновой активности в ЭЭГ выделяют отдельные потенциалы, связанные с какими-либо событиями: вызванные потенциалы, возникающие в ответ на внешние раздражения (слуховые, зрительные и др.); потенциалы, отражающие мозговые процессы при подготовке, осуществлении и окончании отдельных двигательных актов – это «волна ожидания», или условная негативная волна (Уолтер Г., 1966), премоторные, моторные и финальные потенциалы и др. Кроме того, регистрируют различные сверхмедленные колебания длительностью от нескольких секунд до десятков минут (в частности, так называемые «омега-потенциалы» и др.), которые отражают биохимические процессы регуляции функций и психической деятельности.
4. Высшая нервная деятельность
Развивая идеи И. М. Сеченова о рефлекторной основе поведенческой деятельности целостного организма, И. П. Павлов пришел к мысли, что в изменяющихся условиях внешней среды недостаточно обладать стандартными рефлекторными реакциями, а требуется выработка новых рефлексов, адекватных новым условиям существования. Впервые об условных рефлексах он заговорил в известной Мадридской речи в 1903 г.
4.1. Условия образования и разновидности условных рефлексов
Условные рефлексы по многим признакам отличаются от безусловных (табл. 1).
Условные рефлексы у млекопитающих и человека осуществляются корой больших полушарий (в этом также принимают участие таламический отдел промежуточного мозга и в ряде случаев подкорковые ядра).
И. П. Павловым была разработана объективная методика изучения приобретаемых или условных рефлексов, которая основывалась на изоляции обследуемого организма от посторонних раздражений и на точной регистрации сигнала и ответа на него. Исследования проводились на собаках в звукоизолированных камерах («башнях молчания»), куда подавались дозированные раздражения светом, звуком, механическими раздражениями кожи и пр. В качестве ответной реакции было выбрано выделение слюны, которая отводилась от одного из слюнных протоков, выведенного на наружную поверхность щеки (методика фистулы слюнной железы).
Таблица 1
Различия условных и безусловных рефлексов

В процессе выработки приобретаемых рефлексов должны соблюдаться следующие условия:
• сочетание любого индифферентного раздражителя с каким-либо значимым безусловным раздражением (например, пищевым) – методика безусловного подкрепления;
• индифферентное раздражение должно предшествовать безусловному, чтобы приобрести сигнальное значение;
• нервные центры, к которым адресованы раздражения, должны быть в состоянии оптимального возбуждения.
Например, после предварительного изолированного действия светового сигнала собаке подавалось подкрепление – мясосухарный порошок и регистрировалось выделение слюны. После ряда сочетаний этих сигналов уже одно только включение света вызывало выделение слюны, т. е. был выработан новый рефлекс, биологический смысл которого заключался в подготовке организма к приему пищи.
Механизм образования условного рефлекса заключался в формировании новой рефлекторной дуги, в которой к эфферентной части безусловного рефлекса присоединялась новое афферентное начало рефлекторной дуги, идущее от зрительных путей. Между центрами этих исходных рефлексов сформировалась новая связь, которую И. П. Павлов назвал временной связью, так как в случае прекращения подачи пищи после светового сигнала слюнной условный рефлекс исчезал.
В ходе выработки условного рефлекса наблюдались определенные фазы этого процесса:
1) генерализации (обобщенное восприятие сигнала, когда условная реакция наблюдалась на любой сходный сигнал), основой чего были процессы иррадиации возбуждения в коре больших полушарий;
2) концентрации возбуждения (реакция только на конкретный сигнал), что появлялось за счет вырабатываемого условного торможения на посторонние неподкрепляемые сигналы;
3) стабилизации (упрочения условного рефлекса).
В дальнейших исследованиях условные рефлексы были выработаны в разнообразных экспериментальных условиях (в том числе в условиях свободного поведения) у различных животных, птиц, рыб, черепах, даже у амеб. Изучение биопотенциалов коры больших полушарий показало, что условием образования временной связи между изучаемыми корковыми центрами является пространственная синхронизация их электрической активности.
Различают несколько разновидностей условных рефлексов:
1) натуральные – на сигналы, характеризующие безусловные раздражители (например, запах мяса для слюнного рефлекса), и искусственные – на посторонние сигналы (например, запах мяты);
2) наличные и следовые на условный сигнал, непосредственно предшествующий безусловному подкреплению, и на его следовое влияние;
3) положительные (с активным проявлением ответной реакции) и отрицательные (с ее торможением);
4) условные рефлексы на время – при ритмической подаче условных сигналов ответная реакция появляется через заданный интервал даже при отсутствии очередного сигнала;
5) условные рефлексы первого порядка – на один предшествующий условный раздражитель – и более высоких порядков, когда безусловному подкреплению предшествует сочетание двух последовательно подающихся сигналов (свет + звук) – условный рефлекс второго порядка, трех сигналов (свет + звук + касалка) – условный рефлекс третьего порядка и т. д.
У собак вырабатываются в основном рефлексы третьего порядка, у обезьян – четвертого, у грудного ребенка – 5–6 порядка, у взрослого человека – двадцатого и более порядков. Освоение речи человеком представляет собой формирование огромной цепи условно-безусловных рефлексов, не требующих специального подкрепления.
При формировании новых двигательных навыков возникают особые рефлексы, которые в отличие от сенсорных рефлексов или рефлексов I рода (в которых новой частью рефлекторной дуги была афферентная часть) имеют новую часть рефлекторной дуги в эфферентном отделе (новые исполнительные аппараты – мышцы). Это так называемые инструментальные, или оперантные, рефлексы – рефлексы II рода (Конорский Ю. М., 1970).
4.2. Внешнее и внутреннее торможение условных рефлексов
По своему происхождению торможение условных рефлексов может быть безусловным (врожденным) и условным (выработанным в течение жизни). К безусловному торможению относят охранительное, или запредельное, торможение, возникающее при чрезмерно сильном или длительном раздражении, и внешнее торможение условных рефлексов посторонними для центров условного рефлекса раздражителями (например, нарушение непрочного двигательного навыка у спортсмена в необычных условиях соревнований).
Условное торможение вырабатывается при отсутствии подкрепления условного сигнала. Различают несколько видов условного торможения: угасательное, дифференцировочное и запаздывающее.
Угасание развивается при повторении условного сигнала без подкрепления. Например, имея прочный слюнный условный рефлекс у собаки на вспышку света и затем применяя свет без подкрепления, можно получить последовательно следующие условные ответы – 10, 8, 6, 4, 5, 2, 0, 0, 0 капель слюны.
Дифференцировочное торможение вырабатывается при подкреплении одного условного сигнала (например, звук с частотой 500 Гц) и отсутствии подкрепления сходных с ним сигналов (звук 1000, 200 и 100 Гц), на которые первоначально (в период генерализации условного рефлекса) получался условный ответ. Этот вид торможения, в частности, позволяет спортсмену отдифференцировать сокращения ненужных мышц при выработке двигательного навыка, т. е. имеет важное координационное значение. Процесс воспитания человека сопровождается постоянной дифференцировкой подкрепляемых и осуждаемых обществом поведенческих реакций (что такое «хорошо» и что такое «плохо»).
Запаздывающее торможение формируется при отставлении на определенный отрезок времени подкрепления от условного сигнала. В этом случае сразу после условного сигнала реакция отсутствует (тормозится), но перед моментом подкрепления обнаруживается.
4.3. Динамический стереотип
В жизни обычно встречаются не отдельные условные рефлексы, а сложные их комплексы, в которых они сочетаются с безусловными рефлексами (двигательными, сердечно-сосудистыми, дыхательными и пр.). Систему условных и безусловных рефлексов И. П. Павлов назвал динамическим стереотипом. Она вырабатывается при повторении одного и того же порядка раздражений (ситуаций) и соответственно выражается в цепи закрепленных ответных реакций, т. е. стереотипе. Но изменение внешних условий может вызвать перестройку этой системы или ее разрушение, что отмечается термином «динамический».
Например, у собаки выработан динамический стереотип на определенный порядок из 6 раздражителей, и имеются на них закрепленные условные величины слюноотделения, специфические для каждого сигнала: 1) свет – 12 капель; 2) звук – 20 капель; 3) метроном 120 уд./с – 10 капель; 4) метроном 60 уд./с (неподкрепляемый раздражитель) – 0 капель; 5) свет – 12 капель; 6) звук – 20 капель. Если теперь подавать один и тот же сигнал, то ответная цепь реакций сохранится прежней: 1) свет – 12 капель; 2) свет – 20 капель; 3) свет – 10 капель; 4) свет – 0 капель; 5) свет – 12 капель; 6) свет – 20 капель. Однако изолированное включение светового раздражения сохраняет обычный ответ – 12 капель.
Следовательно, в коре больших полушарий собаки образована цепь последовательно возбуждающихся или затормаживающихся нервных центров, в которой активность каждого автоматически вызывает включение следующего. Подобный стереотип возникает у спортсмена при выработке двигательного навыка, особенно при выполнении стандартных движений. Такой стереотип, связанный с цепью моторных актов, А. Н. Крестовников назвал «двигательный динамический стереотип». Он легче образуется при выполнении циклических упражнений, чем ациклических.
4.4. Типы высшей нервной деятельности, первая и вторая сигнальная система
Случившееся в 1924 г. в Ленинграде сильное наводнение грозило затопить клетки с подопытными собаками, которые испытали сильный стресс. На следующий день обнаружилось, что у некоторых из них пропали прочно выработанные условные рефлексы, но у других рефлексы сохранились. Это навело И. П. Павлова на мысль о различных типах нервной системы у животных. В качестве основных свойств нервной системы И. П. Павлов рассматривал силу возбуждения и торможения, их уравновешенность и подвижность. С учетом этих свойств им были выделены следующие четыре типа высшей нервной деятельности (ВНД), которые оказались сходными с четырьмя темпераментами, выделенными еще Гиппократом в V веке до н. э.
1. Тип сильный неуравновешенный (холерик). Характеризуется сильным процессом возбуждения и более слабым процессом торможения, поэтому легко возбуждается и с трудом затормаживает свои реакции.
2. Тип сильный уравновешенный и высокоподвижный (сангвиник). Отличается сильными уравновешенными и высокоподвижными процессами возбуждения и торможения. Легко переключается с одной формы деятельности на другую, быстро адаптируется к новой ситуации.
3. Тип сильный уравновешенный инертный (флегматик). Имеет сильные и уравновешенные процессы возбуждения и торможения, но малоподвижный – медленно переключающийся с возбуждения на торможение и обратно. С трудом переходит от одного вида деятельности к другому, зато вынослив при длительной работе. Медленно, но прочно адаптируется к необычным условиям внешней среды.
4. Тип слабый (меланхолик). Характеризуется слабыми процессами возбуждения и торможения, с некоторым преобладанием тормозного процесса, мало адаптивен, подвержен неврозам. Зато обладает высокой чувствительностью к слабым раздражениям и может их легко дифференцировать.
Описанные типы имеются у животных и человека. Они представляют собой лишь крайние проявления особенностей нервной системы, между которыми может быть значительное число переходных типов.
Кроме того, И. П. Павлов выделил специфически человеческие типы ВНД, связанные с наличием у человека особой второй сигнальной системы – слова видимого, слышимого, написанного, произносимого, в отличие от первой сигнальной системы, общей для человека и животных, – непосредственных раздражителей внешней или внутренней среды организма. Вторая сигнальная система чрезвычайно расширила адаптационные возможности человека. Ее свойствами являются: обобщение сигналов первой и второй сигнальной системы, появление абстракций (сложных комплексных понятий – мужество, ярость, доброта и пр.), возможность передачи накопленного опыта предшествующих поколений последующим (возникновение науки, культуры и пр.). Вторая сигнальная система таким образом составила основу письменной и устной речи, появления математических и нотных символов, абстрактного мышления человека. Ее деятельность связывают с функциями третичных полей коры больших полушарий, преимущественно левого полушария у правшей, где находятся центры речи.
В связи с различным соотношением у людей реакций, связанных с преобладанием первой или второй сигнальной системы, И. П. Павлов различал специфически человеческие типы нервной системы: «мыслительный» – с преобладанием второй сигнальной системы; «художественный» – с преобладанием первой сигнальной системы. Среди взрослых людей количество лиц с преобладанием второй сигнальной системы составляет около половины населения. Около 25 % составляют лица с преобладанием первой сигнальной системы и примерно 25 % – лица, имеющие равновесие обеих систем. Соответственно этим типам в настоящее время различают две основные формы интеллекта человека: невербальный интеллект, отражающий природные возможности индивида манипулировать с непосредственными (особенно зрительно-пространственными) раздражителями, и вербальный интеллект, отражающий способность манипулировать со словесным материалом, что определяет характер поведенческих реакций, в том числе и в спорте.
5. Нервно-мышечный аппарат
У человека существует три вида мышц: поперечно-полосатые скелетные мышцы, особая поперечно-полосатая сердечная мышца и гладкие мышцы внутренних органов.
5.1. Функциональная организация скелетных мышц
Скелетные мышцы человека содержат около 300 млн мышечных волокон и имеют площадь порядка 3 м2. Целая мышца представляет собой отдельный орган, а мышечное волокно – клетку. Мышцы иннервируются двигательными нервами, передающими из центров моторные команды, чувствительными нервами, несущими в центры информацию о напряжении и движении мышц, и симпатическими нервными волокнами, влияющими на обменные процессы в мышце. Функции скелетных мышц заключаются в перемещении частей тела друг относительно друга, перемещении тела в пространстве (локомоция) и поддержании позы тела.
Функциональной единицей мышцы является двигательная единица, состоящая из мотонейрона спинного мозга, его аксона (двигательного нерва) с многочисленными окончаниями и иннервируемых им мышечных волокон. Возбуждение мотонейрона вызывает одновременное сокращение всех входящих в эту единицу мышечных волокон. Двигательные единицы (ДЕ) небольших мышц содержат малое число мышечных волокон (ДЕ мышц глазного яблока 3–6 волокон, мышц пальцев руки 10–25 волокон), а ДЕ крупных мышц туловища и конечностей – до нескольких тысяч (например, ДЕ икроножной мышцы человека – около 2000 мышечных волокон).
Мелкие мышцы иннервируются из одного сегмента спинного мозга, а крупные мышцы – мотонейронами 2–3 спинальных сегментов. Мотонейроны, иннервирующие одну мышцу, составляют общий мотонейронный пул, в котором могут находиться мотонейроны различных размеров. Большие ДЕ образованы крупными мотонейронами, которые имеют толстые аксоны, множество концевых разветвлений и большое число связанных с ними мышечных волокон. Такие ДЕ имеют низкую возбудимость, генерируют высокую частоту нервных импульсов (порядка 20–50 импульсов в 1 с) и характеризуются высокой скоростью проведения возбуждения. Они включаются в работу лишь при высоких нагрузках на мышцу. Мелкие ДЕ имеют мотонейроны небольших размеров, тонкие и медленно проводящие аксоны, малое число мышечных волокон. Они легко возбудимы и включаются в работу при незначительных мышечных усилиях. Нарастание нагрузки вызывает активацию различных ДЕ скелетной мышцы в соответствии с их размерами – от меньших к большим (правило Хеннемана).
Мышечное волокно представляет собой вытянутую клетку (ее диаметр около 10–100 мкм, а длина 10–12 см). В состав волокна входят его оболочка – сарколемма, жидкое содержимое – саркоплазма, ядро, энергетические центры – митохондрии, белковые депо – рибосомы, сократительные элементы – миофибриллы, а также замкнутая система продольных трубочек и цистерн, расположенных вдоль миофибрилл и содержащих ионы Са2+, – саркоплазматический ретикулум. Поверхностная мембрана клетки через равные промежутки образует поперечные трубочки, входящие внутрь мышечного волокна, по которым внутрь клетки проникает потенциал действия при ее возбуждении.
Миофибриллы – это тонкие волокна (диаметр 1–2 мкм, длина 2–2,5 мкм), содержащие два вида сократительных белков (протофибрилл): тонкие нити актина и вдвое более толстые нити миозина. Они расположены таким образом, что вокруг миозиновых нитей находится 6 актиновых нитей, а вокруг каждой актиновой – 3 миозиновых. Миофибриллы разделены Z-мембранами на отдельные участки – саркомеры, в средней части которых расположены преимущественно миозиновые нити, а актиновые нити прикреплены к Z-мембранам по бокам саркомера. (Разная способность актина и миозина преломлять свет создает в состоянии покоя мышцы ее поперечно-полосатый вид в световом микроскопе.)
Нити актина составляют около 20 % сухого веса миофибрилл. Актин состоит из двух форм белка: 1) глобулярной формы – в виде сферических молекул и 2) палочковидных молекул тропомиозина, скрученных в виде двунитчатых спиралей в длинную цепь. На протяжении этой двойной актиновой нити каждый виток содержит по 14 молекул глобулярного актина (по 7 молекул с обеих сторон), наподобие нитки с бусинками, а также центры связывания ионов Са2+. В этих центрах содержится особый белок (тропонин), участвующий в образовании связи актина с миозином.
Миозин составлен из уложенных параллельно белковых нитей (эта часть представляет собой так называемый легкий меромиозин). На обоих концах его имеются отходящие в стороны шейки с утолщениями – головками (эта часть – тяжелый меромиозин), благодаря которым образуются поперечные мостики между миозином и актином.
5.2. Механизмы сокращения и расслабления мышечного волокна
При произвольной внутренней команде сокращение мышцы человека начинается примерно через 0,05 с (50 мс). За это время моторная команда передается от коры больших полушарий к мотонейронам спинного мозга и по двигательным волокнам к мышце. Подойдя к мышце, процесс возбуждения должен с помощью медиатора преодолеть нервно-мышечный синапс, что занимает примерно 0,5 мс. Медиатором здесь является ацетилхолин, который содержится в синаптических пузырьках в пресинаптической части синапса. Нервный импульс вызывает перемещение синаптических пузырьков к пресинаптической мембране, их опорожнение и выход медиатора в синаптическую щель. Действие ацетилхолина на постсинаптическую мембрану чрезвычайно кратковременно, после чего он разрушается ацетилхолинэстеразой на уксусную кислоту и холин. По мере расходования запасы ацетилхолина постоянно пополняются путем его синтезирования в пресинаптической мембране. Однако при очень частой и длительной импульсации мотонейрона расход ацетилхолина превышает его пополнение, а также снижается чувствительность постсинаптической мембраны к его действию, в результате чего нарушается проведение возбуждения через нервно-мышечный синапс. Эти процессы лежат в основе периферических механизмов утомления при длительной и тяжелой мышечной работе.
Выделившийся в синаптическую щель медиатор прикрепляется к рецепторам постсинаптической мембраны и вызывает в ней явления деполяризации. Небольшое подпороговое раздражение вызывает лишь местное возбуждение небольшой амплитуды – потенциал концевой пластинки (ПКП).
При достаточной частоте нервных импульсов ПКП достигает порогового значения и на мышечной мембране развивается мышечный потенциал действия. Он (со скоростью 5 м/с) распространяется вдоль по поверхности мышечного волокна и заходит в поперечные трубочки внутрь волокна. Повышая проницаемость клеточных мембран, потенциал действия вызывает выход из цистерн и трубочек саркоплазматического ретикулума ионов Са2+, которые проникают в миофибриллы, к центрам связывания этих ионов на молекулах актина.
Под влиянием Са2+ длинные молекулы тропомиозина проворачиваются вдоль оси и скрываются в желобки между сферическими молекулами актина, открывая участки прикрепления головок миозина к актину. Тем самым между актином и миозином образуются так называемые поперечные мостики. При этом головки миозина совершают гребковые движения, обеспечивая скольжение нитей актина вдоль нитей миозина с обоих концов саркомера к его центру, т. е. механическую реакцию мышечного волокна (рис. 10).
Энергия гребкового движения одного мостика производит перемещение на 1 % длины актиновой нити. Для дальнейшего скольжения сократительных белков друг относительно друга мостики между актином и миозином должны распадаться и вновь образовываться на следующем центре связывания Са2+. Такой процесс происходит в результате активации в этот момент молекул миозина. Миозин приобретает свойства фермента АТФазы, который вызывает распад АТФ. Выделившаяся при распаде АТФ энергия приводит к разрушению имеющихся мостиков и образованию в присутствии Са2+ новых мостиков на следующем участке актиновой нити. В результате повторения подобных процессов многократного образования и распада мостиков сокращается длина отдельных саркомеров и всего мышечного волокна в целом. Максимальная концентрация кальция в миофибрилле достигается уже через 3 мс после появления потенциала действия в поперечных трубочках, а максимальное напряжение мышечного волокна – через 20 мс.

Рис. 10. Схема электромеханической связи в мышечном волокне. А – состояние покоя; Б – возбуждение и сокращение; ПД – потенциал действия; ММ – мембрана мышечного волокна; П – поперечные трубочки; Т – продольные трубочки и цистерны с ионами Са2+; а – тонкие нити актина, м – толстые нити миозина с утолщениями (головками) на концах. Зет-мембранами ограничены саркомеры миофибрилл. Толстые стрелки – распространение потенциала действия при возбуждении волокна и перемещение ионов Са2+ из цистерн и продольных трубочек в миофибриллы, где они содействуют образованию мостиков между нитями актина и миозина и скольжение этих нитей (сокращение волокна) за счет гребковых движений головок миозина
Весь процесс от появления мышечного потенциала действия до сокращения мышечного волокна называется электромеханической связью (или электромеханическим сопряжением). В результате сокращения мышечного волокна актин и миозин более равномерно распределяются внутри саркомера, и исчезает видимая под микроскопом поперечная исчерченность мышцы.
Расслабление мышечного волокна связано с работой особого механизма – «кальциевого насоса», который обеспечивает откачку ионов Са2+ из миофибрилл обратно в трубочки саркоплазматического ретикулума. На это также тратится энергия АТФ.
5.3. Одиночное и тетаническое сокращение. Электромиограмма
При единичном надпороговом раздражении двигательного нерва или самой мышцы возбуждение мышечного волокна сопровождается одиночным сокращением. Эта форма механической реакции состоит из трех фаз: латентного, или скрытого, периода; фазы сокращения; фазы расслабления. Самой короткой фазой является скрытый период, когда в мышце происходит электромеханическая передача. Фаза расслабления обычно в 1,5–2 раза более продолжительна, чем фаза сокращения, а при утомлении затягивается на значительное время.
Если интервалы между нервными импульсами короче, чем длительность одиночного сокращения, то возникает явление суперпозиции – наложение механических эффектов мышечного волокна друг на друга и наблюдается сложная форма сокращения – тетанус. Различают две формы тетануса – зубчатый тетанус, возникающий при более редком раздражении, когда происходит попадание каждого следующего нервного импульса в фазу расслабления отдельных одиночных сокращений, и сплошной, или гладкий, тетанус, возникающий при более частом раздражении, когда каждый следующий импульс попадает в фазу сокращения (рис. 11). Таким образом (в некоторых границах) между частотой импульсов возбуждения и амплитудой сокращения волокон ДЕ существует определенное соотношение: при небольшой частоте (например, 5–8 имп. в 1 с) возникают одиночные сокращения, при увеличении частоты (15–20 имп. в 1 с) – зубчатый тетанус, при дальнейшем нарастании частоты (25–60 имп. в 1 с) – гладкий тетанус. Одиночное сокращение – более слабое и менее утомительное, чем тетаническое. Зато тетанус обеспечивает в несколько раз более мощное, хотя и кратковременное сокращение мышечного волокна.

Рис. 11. Одиночное сокращение, зубчатый и сплошной тетанус камбаловидной мышцы человека (по: Н. В. Зимкин и др., 1984); верхняя кривая – сокращение мышцы, нижняя – отметка раздражения мышцы, справа указана частота раздражения
Сокращение целой мышцы зависит от формы сокращения отдельных ДЕ и их координации во времени. При обеспечении длительной, но не очень интенсивной работы, отдельные ДЕ сокращаются попеременно (рис. 12), поддерживая общее напряжение мышцы на заданном уровне (например, при беге на длинные и сверхдлинные дистанции). При этом отдельные ДЕ могут развивать как одиночные, так и тетанические сокращения, что зависит от частоты нервных импульсов. Утомление в этом случае развивается медленно, так как, работая по очереди, ДЕ в промежутках между активацией успевают восстанавливаться. Однако для мощного кратковременного усилия (например, поднятия штанги) требуется синхронизация активности отдельных ДЕ, т. е. одновременное возбуждение практически всех ДЕ, что, в свою очередь, требует одновременной активации соответствующих нервных центров и достигается в результате длительной тренировки. При этом осуществляется мощное и весьма утомительное тетаническое сокращение.

Рис. 12. Различные режимы работы двигательных единиц (ДЕ)
Амплитуда сокращения одиночного волокна не зависит от силы надпорогового раздражения (закон «Все или ничего»). В отличие от этого, при нарастании силы надпорогового раздражения сокращение целой мышцы постепенно растет до максимальной амплитуды.
Работа мышцы с небольшой нагрузкой сопровождается редкой частотой нервных импульсов и вовлечением небольшого числа ДЕ. В этих условиях, накладывая отводящие электроды на кожу над мышцей и используя усилительную аппаратуру, можно на экране осциллографа или с применением чернильной записи на бумаге зарегистрировать одиночные потенциалы действия отдельных ДЕ. В случае же значительных напряжений потенциалы действия многих ДЕ алгебраически суммируются, возникает сложная интегрированная кривая записи электрической активности целой мышцы – электромиограмма (ЭМГ).
Форма ЭМГ отражает характер работы мышцы: при статических усилиях она имеет непрерывный вид, а при динамической работе – вид отдельных пачек импульсов, приуроченных в основном к начальному моменту сокращения мышцы и разделенных периодами «электрического молчания». Особенно хорошо ритмичность появления подобных пачек наблюдается у спортсменов при циклической работе (рис. 13). У маленьких детей и неадаптированных к такой работе лиц четких периодов отдыха не наблюдается, что отражает недостаточное расслабление мышечных волокон работающей мышцы.
Рис. 13. Электромиограмма мышц-антагонистов при циклической работе
Чем больше внешняя нагрузка и сила сокращения мышцы, тем выше амплитуда ее ЭМГ. Это связано с увеличением частоты нервных импульсов, вовлечением большего числа ДЕ в мышце и синхронизацией их активности. Современная многоканальная аппаратура позволяет производить одновременную регистрацию ЭМГ многих мышц на разных каналах. При выполнении спортсменом сложных движений можно видеть на полученных ЭМГ кривых не только характер активности отдельных мышц, но и оценить моменты и порядок их включения или выключения в различные фазы двигательных актов. Записи ЭМГ, полученные в естественных условиях двигательной деятельности, можно передавать к регистрирующей аппаратуре по телефону или радиотелеметрически. Анализ частоты, амплитуды и формы ЭМГ (например, с помощью специальных компьютерных программ) позволяет получить важную информацию об особенностях техники выполняемого спортивного упражнения и степени ее освоения обследуемым спортсменом.
По мере развития утомления при той же величине мышечного усилия амплитуда ЭМГ нарастает. Это связано с тем, что снижение сократительной способности утомленных ДЕ компенсируется нервными центрами вовлечением в работу дополнительных ДЕ, т. е. путем увеличения количества активных мышечных волокон. Кроме того, усиливается синхронизация активности ДЕ, что также повышает амплитуду суммарной ЭМГ.
5.4. Монофункциональные основы мышечной силы
Движение является результатом взаимодействия внутренних и внешних сил, развиваемых в опорно-двигательном аппарате, – активных (возникающих при сокращении или напряжении мышцы во время ее возбуждения) и пассивных (упругое напряжение при растяжении мышцы, сопротивление мышцы и ее сухожилия).
Сила мышцы зависит от ряда морфологических и физиологических факторов: количества и свойств мышечных волокон в мышце, исходной длины мышцы, характера нервных импульсов, механических условий действия мышцы на кости скелета.
Сила мышцы является суммой силы отдельных ее мышечных волокон. Подсчитано, что 1 одиночное мышечное волокно икроножной мышцы развивает напряжение 100–200 мг, 1 ДЕ икроножной мышцы человека содержит около 2000 мышечных волокон и развивает напряжение 200–400 г, 1 икроножная мышца содержит около 1000 ДЕ и развивает напряжение 200–400 кг.
Большое значение имеет анатомическое строение мышцы. В параллельно-волокнистых и веретенообразных мышцах (камбаловидная мышца и др.) сила мышц тем больше, чем больше ее анатомический поперечник, т. е. площадь поперечного сечения целой мышцы. В перистых мышцах (двуглавая мышца и др.) физиологический поперечник, т. е. площадь поперечного сечения всех мышечных волокон, гораздо больше, чем ее анатомический поперечник. В такой мышце упаковано значительно больше мышечных волокон и соответственно больше ее сила.
На силу сокращения мышцы влияет ее исходная длина, так как от нее зависит возможное количество поперечных мостиков между актином и миозином. Предполагают, что в каждом цикле присоединения-отсоединения поперечных мостиков расходуется энергия 1 молекулы АТФ на 1 поперечный мостик. Следовательно, чем больше образуется в мышечном волокне актиномиозиновых мостиков, тем выше скорость расщепления АТФ, больше тяга сократительных белков и соответственно больше развиваемая мышцей сила.
Наибольшее количество актиномиозиновых контактов образуется при небольшом растяжении мышцы до некоторой оптимальной длины. При значительном растяжении саркомера нити актина далеко расходятся в стороны и практически не контактируют с расположенным в средней части саркомера миозином. В случае же резкого уменьшения длины саркомера нити актина в центре перекрывают друг друга, препятствуя контактам с миозином и также уменьшая число образуемых мостиков. В связи с этими особенностями взаимодействия сократительных белков наибольшая сила мышцы проявляется при некотором ее предварительном растяжении.